FunctionEdit
ABA:n uskottiin alun perin osallistuvan abskissioon, mistä se sai nimensä. Nykyään tiedetään, että näin on vain harvoilla kasveilla. ABA-välitteisellä signaloinnilla on myös tärkeä rooli kasvien vasteissa ympäristöstressiin ja kasvipatogeeneihin. ABA:n biosynteesin kasvigeenit ja reitin kulku on selvitetty. ABA:ta tuottavat myös eräät kasvipatogeeniset sienet eri biosynteesireitin kautta kuin ABA:n biosynteesi kasveissa.
Talveen valmistauduttaessa ABA:ta tuotetaan päätelehtisilmuissa. Tämä hidastaa kasvin kasvua ja ohjaa lehtien alkusilmuja kehittämään suomuja suojaamaan lepotilassa olevia silmuja kylmän kauden aikana. ABA estää myös solujen jakautumista verisuonikambiumissa, jolloin sopeudutaan talven kylmiin olosuhteisiin keskeyttämällä primaari- ja sekundaarikasvu.
Absisiinihappoa tuotetaan myös juurissa vasteena maaperän vesipotentiaalin alenemiseen (joka liittyy kuivaan maaperään) ja muihin tilanteisiin, joissa kasvi voi olla stressissä. ABA siirtyy sitten lehtiin, jossa se muuttaa nopeasti stomataalisten suojasolujen osmoottista potentiaalia, jolloin ne kutistuvat ja stomata sulkeutuu. ABA:n aiheuttama stomataalien sulkeutuminen vähentää transpiraatiota (veden haihtumista stomataalien ulkopuolelle), mikä estää veden lisäkadon lehdistä silloin, kun vettä on vähän saatavilla. Lehtien ABA-pitoisuuden ja lehtien konduktanssin (stomaattiresistanssi) välillä havaittiin läheinen lineaarinen korrelaatio lehtipinta-alan perusteella.
Siementen itämistä estää ABA antagonismissa gibberelliinin kanssa. ABA estää myös siementen lepotilan häviämisen.
Nottinghamin Arabidopsis Stock Centre on tunnistanut useita ABA-mutantteja Arabidopsis thaliana -kasveja, joita on saatavilla Nottinghamin Arabidopsis Stock Centrestä – sekä sellaisia, joilla on ABA:n tuotantopuutos, että sellaisia, joiden herkkyys ABA:n vaikutukselle on muuttunut. Kasvit, jotka ovat yli- tai aliherkkiä ABA:lle, osoittavat fenotyyppejä siementen horroksessa, itämisessä ja stomataalien säätelyssä, ja joillakin mutanteilla esiintyy kitukasvuisuutta ja ruskeita/keltaisia lehtiä. Nämä mutantit heijastavat ABA:n merkitystä siementen itämisessä ja alkion varhaisessa kehityksessä.
Pyrabaktiini (pyridyyliä sisältävä ABA:n aktivaattori) on naftaleenisulfonaamidi hypokotyylisolujen laajenemisen estäjä, joka on siementen ABA-signalointireitin agonisti. Se on ensimmäinen ABA-reitin agonisti, joka ei ole rakenteellisesti sukua ABA:lle.
HomeostaasiEdit
BiosynteesiEdit
Absisiinihappo (ABA) on isoprenoidinen kasvihormoni, joka syntetisoituu plastidaalisessa 2-C-metyyli-D-erytritoli-4-fosfaatti (MEP) -reitissä; Toisin kuin rakenteellisesti samankaltaiset seskviterpeenit, jotka muodostuvat mevalonihaposta peräisin olevasta esiasteesta farnesyylidifosfaatista (FDP), ABA:n C15-runko muodostuu MEP:n C40-karotenoidien pilkkomisen jälkeen. Zeaksantiini on ensimmäinen ABA:n esiaste; entsyymikatalysoitujen epoksidaatioiden ja isomerisaatioiden sarja violaksantiinin kautta ja C40-karotenoidin lopullinen pilkkominen dioksihapetusreaktiolla tuottavat ABA:n proksimaalisen esiasteen, ksantoksiinin, joka hapetetaan edelleen ABA:ksi absisialdehydin kautta.

Abamiini on suunniteltu, syntetisoitu, kehitetty ja sitten patentoitu ensimmäisenä spesifisenä ABA:n biosynteesin estäjänä, joka mahdollistaa ABA:n endogeenisten pitoisuuksien säätelyn.
ABA:n biosynteesin sijoittuminen ja ajoitus Muokkaa
- Vapautuu kasvullisen kudoksen kuivumisen aikana ja juurten kohdatessa maaperän tiivistymisen.
- Syntetisoituu vihreissä hedelmissä talvikauden alussa
- Syntetisoituu kypsyvissä siemenissä vakiinnuttaen horroksen
- Mobilisoituu lehden sisällä ja voi siirtyä nopeasti juurista lehtiin ksyleemin transpiraatiovirtauksen kautta
- Tuotetaan vasteena ympäristöstressille, kuten lämpöstressille, vesistressille, suolastressille
- Syntetisoituu kaikissa kasvinosissa, esim, juuret, kukat, lehdet ja varret
- ABA:ta syntetisoidaan lähes kaikissa soluissa, joissa on kloroplasteja tai amyloplasteja
InaktivointiEdit
ABA voidaan katabolisoida faasihapoksi CYP707A:n (ryhmä P450-entsyymejä) välityksellä tai inaktivoida glukoosikonjugaatiolla (ABA-glukoosiesteri) AOG-entsyymin välityksellä. CYP707A:n kautta tapahtuva katabolia on erittäin tärkeää ABA:n homeostaasin kannalta, ja näiden geenien mutantit keräävät yleensä suurempia ABA-pitoisuuksia kuin ABA:n biosynteettisiä geenejä yliekspressoivat linjat. Maaperän bakteereissa on raportoitu vaihtoehtoinen katabolinen reitti, joka johtaa dehydrovomifoliolioliin vomifoliolidehydrogenaasientsyymin välityksellä.
VaikutuksetEdit
- Antitranspirantti – Indusoi stomataalien sulkeutumista, mikä vähentää transpiraatiota estääkseen vesihävikin.
- Estä hedelmien kypsymistä
- Vastaa siementen lepotilaan estämällä solujen kasvua – estää siementen itämistä
- Estä kinetiininukleotidin synteesiä
- Alasäätelee fotosynteesiin tarvittavia entsyymejä.
- Vaikuttaa endodermikseen estäen juurten kasvua, kun ne altistuvat suolaisille olosuhteille
- Hidastaa solujen jakautumista
- Dormancy inducer – Sitä käytetään siementen horroksen indusoimiseen .
- käytetään anti – transpirantteina – Kuivuusalttiilla alueilla , vesistressi on vakava ongelma maataloustuotannossa. joten ehdotetaan ABA:n ruiskutuksia, jotka aiheuttavat osittaisen stomata sulkeutumisen muutamaksi päiväksi , vähentääkseen transpiratiivista vesihäviötä
SignaalikaskadiEdit
>
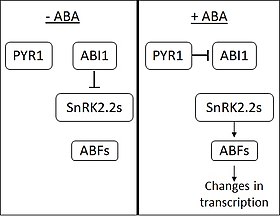
Ababan puuttuessa, fosfataasi ABI1-INSENSITIVE1 (ABI1) estää SNF1:een liittyvien proteiinikinaasien (alaryhmä 2) (SnRK2s) toimintaa. ABA:ta havaitsevat PYRABACTIN RESISTANCE 1 (PYR1) ja PYR1:n kaltaiset kalvoproteiinit. ABA:n sitoutuessa PYR1 sitoutuu ABI1:een ja estää sen toimintaa. Kun SnRK2:t vapautuvat inhibitiosta, ne aktivoivat useita ABA RESPONSIVE ELEMENT-BINDING FACTOR (ABF) -perheeseen kuuluvia transkriptiotekijöitä. ABF:t aiheuttavat sitten muutoksia useiden geenien ilmentymisessä. ABA:n uskotaan säätelevän noin 10 % kasvigeeneistä.