- Introduction
- Výsledky
- Akumulace buněk a jejich přichycení k minerálu vyžaduje geny pro chemotaxi a extracelulární transport elektronů u S. oneidensis MR-1
- Characterizing Congregation around IEA in Other Shewanella Strains
- Diskuse
- Materiál a metody
- Kultivace a kmeny
- Syntéza minerálů
- Časosběrné experimenty s akumulací buněk po fotobuňkách (CAAP)
- Mikroskopické zachycení pohybu buněk
- Analýza plavání
- Příspěvky autorů
- Prohlášení o střetu zájmů
- Poděkování
- Doplňkový materiál
Introduction
Na konci 80. let 20. století byla objevena Shewanella oneidensis MR-1 (Myers a Nealson, 1988a) a později několik druhů Geobacter (Lovley et al., 1993; Champine et al., 2000) bylo prokázáno, že jsou schopny přenášet elektrony na nerozpustné akceptory elektronů (IEA), jako jsou nerozpustné oxidy kovů a/nebo nabité elektrody: proces se nazývá extracelulární transport elektronů (EET) (Myers a Nealson, 1988b; Venkateswaran a kol., 1999; Bond a Lovley, 2003). Tato schopnost vzbudila značný zájem s ohledem na biogeochemický cyklus, bioremediaci, korozi, zpracování nanomateriálů a výrobu energie (Bretschger a kol., 2007; Kan a kol., 2011; Hsu a kol., 2012). Přestože je známo několik skupin mikrobů schopných EET, hlavní mechanistické studie byly provedeny pouze se dvěma modelovými systémy, Shewanella (Fredrickson et al., 2008; Shi et al., 2009) a Geobacter (Lovley et al.,
Dosud bylo izolováno více než 100 dalších kmenů Shewanella z nejrůznějších stanovišť včetně otevřeného vodního sloupce, pískovcových břidlic, mořských a sladkovodních sedimentů, ropovodů, ropných solanek a dokonce i společenstev řas na vrcholu antarktického ledu (Hau a Gralnick, 2007). Genomy více než 20 těchto druhů byly plně sekvenovány (Fredrickson et al., 2008). U několika z těchto druhů byla prokázána schopnost EET na IEA, včetně S. oneidensis MR-1, Shewanella putrefaciens CN32, S. sp. ANA-3, S. sp. W3-18-1, Shewanella amazonensis SB2B, Shewanella frigidimarina NCIMB 400 a Shewanella loihica PV-4 (Fredrickson et al., 1998; Venkateswaran et al., 1998; Gao et al., 2006; Bretschger, 2008). Ostatní zástupci rodu Shewanella, jako Shewanella baltica OS217 a Shewanella denitrificans OS155 (tabulka 1), nejsou schopni EET (Brettar et al., 2002). Ze všech těchto kmenů bylo shlukování v reakci na IEA studováno pouze u MR-1.

Tabulka 1. Genetické srovnání Shewanella spp. a jejich původních stanovišť.
V rámci rodu Shewanella byl mechanismus EET u MR-1 charakterizován nejrozsáhleji. MR-1 využívá několik přístupů k redukci nerozpustných IEA: (1) přímá EET prostřednictvím extracelulárních multihemových cytochromů (Beliaev a Saffarini, 1998; Myers a Myers, 2001, 2002; Meyer a kol., 2004; Mitchell a kol., 2012; Kracke a kol., 2015). (obr. 1A); (2) zprostředkované EET pomocí rozpustných raketoplánů vázaných na membránové cytochromy (Lovley et al., 1996; Marsili et al., 2008; Li et al., 2012; Kotloski a Gralnick, 2013; Okamoto et al., 2014); (3) zprostředkovaná EET využívající vodivá rozšíření vnější membrány, která obsahují cytochromy (Gorby et al., 2006; El-Naggar et al., 2010); a (4) vodivé extracelulární matrice obsahující vodivé a polovodivé minerály (Kato et al., 2010). U kmene MR-1 bylo identifikováno několik genů, u nichž se ukázalo, že jsou pro EET nezbytné (obr. 1A a tab. 2), včetně tetrahemového cytochromu cymA (SO_4591) a kombinace mtrBC/omcA (SO_1776, SO_1778 a SO_1779), které kódují dekahemovou složku cytochromu c a tetrahemový cytochrom c nezbytný pro redukci několika anaerobních akceptorů elektronů, včetně oxidů kovů (Myers a Myers, 2001, 2002; Schwalb a kol., 2003). Protože všechny tyto mechanismy jsou závislé na blízkosti buňky k IEA pro EET, je důležité porozumět vnímání buňky a čisté plavecké migraci směrem k IEA. Pokud jde o shromažďování, redoxní taxis nebo energetický taxis, bylo provedeno mnoho studií na MR-1 vzhledem k jeho všestrannému využití elektronových akceptorů (Bencharit a Ward, 2005; Baraquet a kol., 2009; Harris a kol., 2010). Energetická taxis je termín, který široce zahrnuje aerotaxi, fototaxi, redoxní taxis, taxis k alternativním akceptorům elektronů a chemotaxi k oxidovatelným substrátům (Alexandre et al., 2004).
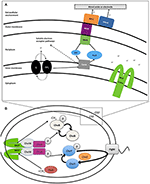
Obrázek 1. Taxis k alternativním akceptorům elektronů. (A) Extracelulární transport elektronů (EET) u Shewanella oneidensis MR-1 buduje protonovou hybnou sílu (pmf). Za anaerobních podmínek mohou příslušníci rodu Shewanella transportovat elektrony z vnitřní membrány, periplazmy, vnější membrány a elektrody prostřednictvím řetězce cytochromů a menachinonů (MQ) známého pod souhrnným názvem extracelulární elektronový transport (EET). Exprese cytochromů EET může kolísat v závislosti na dostupných akceptorech elektronů v prostředí. Počet hemových skupin typu c obsahujících železo je uveden uvnitř (v závorce). Elektrony proudí od donorů elektronů, jako je laktát, k redukci chinonů (Q), které současně přenášejí H+ do periplazmy, budují pmf a zároveň předávají elektrony CymA. V případě nepřítomnosti rozpustných akceptorů elektronů jsou elektrony přenášeny na komplex vnější membrány MtrCAB. Tento komplex MtrCAB může odevzdávat elektrony přímo terminálnímu akceptoru elektronů, a to buď minerální nebo anodové elektrodě, nebo prostřednictvím molekul flavinu. pmf pohání produkci ATP a rotaci polárních bičíků. „Self-sensing“ methyl-accepting chemotaxis proteins (MCPs) řídí rotaci bičíků prostřednictvím chemotaxis signal transduction system (B) a mohou detekovat změny koncentrace H+ během redukce kovů. (B) Pravděpodobná dráha přenosu signálu chemotaxe u Shewanella. V reakci na stimulaci se struktura MCP posouvá jako píst, což způsobuje zpomalení nebo zastavení autofosforylace CheA. CheY a CheB proto nejsou fosforylovány a tento nedostatek CheY-P umožňuje plynulé plavání. Tato stimulace má také další účinek – CheB je bez fosforylace neaktivní (nemůže fungovat jako methylesteráza) a to umožňuje proteinu CheR (neustále aktivní methyltransferáze) předbíhat a volně methylovat dimerní methyl-akceptující oblast MCP (doména HAMP). Tato metylace MCP působí na zvýšení rychlosti autofosforylace histidinového zbytku CheA. Proto má systém přenosu signálu kontrolu nad frekvencí bičíků při zvýšeném nebo sníženém množství podnětů, což vede k sérii plavání „run-and-reversal“.

Tabulka 2. Geny MR-1 popsané v textu.
Je dobře zdokumentováno, že akumulace buněk MR-1 v reakci na rozpustné akceptory elektronů je formou energetické taxe, která závisí na toku H+ a vytvoření protonové hybné síly (Baraquet et al., 2009) (obr. 1). Součástí této odpovědi je i rychlejší plavání, které je pozorováno také u elektronových akceptorů, jako je riboflavin nebo antrachinon 2,6-disulfonát (AQDS) (Bencharit a Ward, 2005; Harris a kol., 2010; Li a kol., 2012). Naproti tomu hromadění buněk kolem IEA, které bylo nazváno kongruence (Nealson et al., 1995), zahrnuje jak zvýšenou rychlost plavání při kontaktu s IEA (tzv. elektrokineze), tak zvýšený počet plaveckých obratů při poklesu PMF. Plavecké reverzace umožňují vícenásobná přechodná setkání buněk s IEA (trvající 1-100 ms) a rychlost, s jakou plavecké buňky během kongregace přecházejí k ireverzibilnímu přichycení k IEA, nebyla dosud kvantifikována.
Mezinformace, kterou buňky využívají k lokalizaci IEA, zůstává nejasná (Nealson et al., 1995; Bencharit a Ward, 2005; Harris et al., 2010). Dřívější zprávy navrhovaly, že jiný studovaný modelový organismus Geobacter se hromadí v okolí IEA tím, že vnímá gradient redukovaných kovových iontů (Childers et al., 2002), avšak redukované kovové ionty nejsou zapojeny do mechanismu vnímání, který používá MR-1 (Bencharit a Ward, 2005). Nedávno bylo popsáno shromáždění bakterií v reakci na poizované elektrody, což ukazuje spíše na redoxní snímání než na snímání kovových iontů jako spouštěč reakce (Harris et al., 2010). Kladné potenciály přiložené k elektrodě (200-600 mV oproti Ag/AgCl) způsobily shlukování MR-1 podobně jako v případě oxidů kovů (Harris et al., 2012). Redoxní potenciály minerálů obsahujících MnO2 se pohybují mezi 400 a 600 mV vs. Ag/AgCl a přijímají elektrony ochotněji než minerály Fe(OH)3, které nesou ekvivalentní poizovaný potenciál 100 až 300 mV vs. Ag/AgCl (Burdige, 1993). V předchozí studii se předpokládalo, že charakteristické plavání MR-1 kolem IEA je regulováno dvěma samočinnými receptory chemotaxe, methyl-akceptujícími proteiny (MCP) s, Ca+ kanálovým receptorem chemotaxe (Cache) doménou (SO_2240) a Per/Arnt/Sim (PAS) doménami (SO_1385), a proteinkinázou CheA-3 (SO_3207), která přenáší signál chemotaxe, aby buňka mohla odebírat vzorky redoxního potenciálu nebo schopnosti povrchu přijímat elektrony (tabulka 2 a obrázek 1B) (Harris et al., 2012). Před touto studií nebyly známy geny zodpovědné za motilitou řízené přichycení buněk na IEA a v jejím okolí.
V této zprávě studujeme mechanismus shromažďování různých kmenů Shewanella sledováním vzorců plavání a přichycení buněk na MnO2 a Fe(OH)3 . Kromě screeningu WT a několika delečních mutantů MR-1 byly prověřeny i další druhy rodu Shewanella (viz tabulka 1). Ukázalo se, že mnoho kmenů je schopno shlukovat se kolem MnO2 i FeOH3, zatímco jiné reagují selektivně na MnO2 (SB2B) nebo Fe(OH)3 (W3-18-1). Při sledování přilnavosti buněk k minerálním povrchům jsme pozorovali silnou korelaci mezi schopností buněk shlukovat se a přilnavostí buněk k povrchu IEA. Poté jsme porovnali genomy těchto druhů, abychom našli kandidátní geny podílející se na fenotypech plavání ve shlucích, akumulaci a přichycení buněk v reakci na specifický povrch IEA.
Výsledky
Akumulace buněk a jejich přichycení k minerálu vyžaduje geny pro chemotaxi a extracelulární transport elektronů u S. oneidensis MR-1
Většina prvních studií energetické taxisace u MR-1 využívala metodu testů swarm plate (Nealson et al., 1995; Baraquet et al., 2009; Li et al., 2012). Zde nabízíme hlubší charakterizaci těchto dosud neznámých mechanismů energetické taxis pomocí kvantitativní metody sledování plavání jednotlivých buněk tekutým médiem v reakci na IEA a akumulace buněk po fotobělení (CAAP) k měření rychlosti přichycení buněk (Li et al., 2010). CAAP využívá ozáření UV laserem z konfokálního mikroskopu k nevratnému ztmavení (zhasnutí fluorescence) buněk označených GFP na dané IEA a v jejím okolí. Kvantifikací fluoreskujících buněk při jejich pohybu do ztmavené zóny lze kvantifikovat rychlost akumulace a uchycení a porovnat různé kmeny. Aby bylo možné považovat pohyblivou buňku za přichycenou, ověří se její poloha analýzou 3D obrazu zorného pole. Pokud buňka spočine na povrchu minerálu, považuje se za „připojenou“. Síla vazeb mezi nehybnými buňkami MR-1 a povrchy byla měřena pomocí optické pinzety v samostatné studii (Gross a El-Naggar, 2015). Tato metoda umožňuje rozlišit mezi cíleným přichycením buněk a náhodným elektrostatickým přichycením.
Jak ukazuje obrázek 2, buňky WT MR-1 se přichytily k povrchu minerálu po 30 minutách s určitou velkou odchylkou v počtu, 2 655 ± 1 352 buněk/mm2. Během této doby buňky vykazují „touch and go“ plavání a navazují přechodný kontakt s minerálním povrchem MnO2. Po 2 h se počet nových buněk přichycených k povrchu minerálu zvýšil na 4 300 ± 584 buněk/mm2 povrchu minerálu, zatímco další pohyblivé buňky se nadále shromažďovaly (videa S1, S2 a S17). Mutanti s trojitou delecí klíčových genů EET mtrBC/omcA (SO_1776, SO_1778, SO_1779), jednoduchou delecí cymA (SO_4591) a hlavního energetického taxonu chemoreceptoru MCP Cache (SO_2240) byli již dříve spojeni s fenotypem shlukování při plavání (Harris et al., 2012). Deleční mutanti (ΔcymA, Δmcp_cache a ΔmtrBC/omcA) byli všichni pohybliví a schopní měnit směr plavání, ale nebyli schopni shlukování a během experimentu vykazovali malou nebo žádnou vazbu na MnO2 (obr. 2C a obr. S1A,B). Akumulace WT v odbarvené zóně v t = 2 h činí v průměru 4 300 ± 584 buněk/mm2, zatímco mutanti Δmcp_cache a ΔmtrBC/omcA se hromadí v zanedbatelném počtu (Obrázek 2C a S1CD).
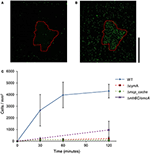
Obrázek 2. Plovoucí MR-1 může migrovat k nerozpustným minerálům akceptorů elektronů a připojovat se. Reprezentativní snímek konfokální fluorescenční mikroskopie buněk WT MR-1 při t = 0 a t = 120 min (A,B). Plovoucí buňky WT MR-1 GFP byly zavedeny k částicím MnO2 (červený tečkovaný obrys) v anaerobní uzavřené kapiláře. V čase t = 0 byly všechny buňky nevratně fotoběleny v oblasti 250 x 250 μm kolem částice (A). Poté byly spočítány fluoreskující buňky mimo vybělenou zónu, které vpluly do rámečku a přilnuly k povrchu minerálu. Černá svislá stupnice vpravo představuje 100 μm. Graf (C) porovnává přichycení WT MR-1 na částici MnO2 po dobu 120 minut s chemotaxí a mutanty s delecí extracelulárního přenosu elektronů (MR-1 Δmcp_cache, ΔcymA a ΔmtrBC/omcA). Chybové úsečky zahrnují 2 std odchylky.
Characterizing Congregation around IEA in Other Shewanella Strains
Devět kmenů Shewanella bylo testováno na schopnost shlukovat se kolem minerálních povrchů MnO2 a Fe(OH)3 pomocí časové řady testů přichycení buněk k minerálu (tabulka 3 a videa S3-S14). Stejně jako u MR-1 mělo všech devět zkoumaných kmenů jeden polární bičík a změna směru plavání byla provedena obrácením rotace bičíku (údaje nejsou uvedeny). Plavecké dráhy v rámci jednoho experimentu byly roztříděny do dvou samostatných skupin na základě dráhy plavání (tabulka 3): ty, které se dotkly povrchu nerozpustného oxidu kovu (plavaly do 2 μm), ve srovnání s těmi, které se nedotkly (plavaly >2 μm). Plavci, kteří se dostali do kontaktu a prokázali významné zvýšení frekvence couvání a rychlosti plavání oproti skupině, která se do kontaktu nedostala (P < 0,05), jsou klasifikováni jako pozitivní pro chování při sběru, jak je označeno písmeny s horním indexem (Tabulka 3).

Tabulka 3. Rychlost plavání bakterií a frekvence reverze v okolí minerálů oxidů kovů.
Kmeny ANA3, CN32 a PV-4 byly kromě MR-1 pozitivní pro shlukování v okolí MnO2 i Fe(OH)3 (tabulka 3, obrázek 3; videa S1-S8). Vykazovaly maximální míru reverze, když se nacházely mezi 5 a 40 μm od povrchu částic MnO2 nebo FeOH3 (obrázek 3). Tři kmeny: OS217, NCIMB400 a OS155 nebyly v čase t = 30 min pohyblivé v reakci na minerály Fe(OH)3 nebo MnO2 (tabulka 3). Například frekvence reverzace plovoucích buněk MR-1, které přišly do kontaktu s MnO2, byla 0,94 ± 0,53 reverzace/s, zatímco frekvence reverzace nekontaktních buněk byla 0,62 ± 0,73 reverzace/s. Rychlost se také zvýšila ve skupině MR-1, která se dostala do kontaktu, z 19,26 ± 11,2 μm/s ve skupině, která se nedostala do kontaktu, na 24,37 ± 6 μm/s ve skupině, která se dostala do kontaktu. Protože došlo jak ke statisticky významnému zvýšení rychlosti plavání, tak ke zpětnému pohybu tak, aby buňky mohly zůstat v blízkosti částice oxidu kovu, byl tento kmen označen za kongruenčně pozitivní.
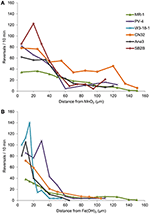
Obrázek 3. (A,B) Reverzní frekvence (osa y) plovoucích buněk rodu Shewanella v závislosti na vzdálenosti (osa x) od částice IEA. Průměrné reverzní frekvence jednotlivých sledovaných plovoucích buněk jsou rozděleny do binů po 5 μm podél osy x pro experimenty s částicí MnO2 (A) nebo Fe(OH)3 (B).
Plavecké dráhy Shewanella spp (30 s), které prokázaly preferenci minerálů oxidů kovů, jsou zvýrazněny na obrázcích 4A-F. Na obrázku SB2B buňky nevykazovaly žádnou plaveckou reakci na Fe(OH)3 (obrázek 4D), zatímco vykazovaly aktivní shlukování kolem částic MnO2 (obrázek 4A). Naproti tomu buňky W3-18-1 se shromažďovaly kolem částic Fe(OH)3 (Obrázek 4C), ale vykazovaly výrazně sníženou plaveckou a reverzní aktivitu kolem MnO2 (Obrázek 4F; videa S9, S14).
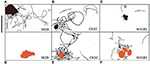
Obrázek 4. (A-F) Panel tří plaveckých drah (30 s) Shewanella spp. prokázal preferenci minerálů oxidů kovů. Bylo studováno plavecké chování tří pohyblivých Shewanella spp. v reakci na MnO2 (horní řádek-černě zbarvená částice) a Fe(OH)3 (spodní řádek-oranžově zbarvená částice). Ke shlukování dochází za anaerobních podmínek, plavecké dráhy jsou znázorněny černě. SB2B (vlevo), CN32 (uprostřed) a W3181 (vpravo). Plavání buněk bylo sledováno ručně. Měřítko vlevo nahoře = 50 μm. Hnědý barevný tvar představuje MnO2 a oranžový Fe(OH)3. Pokud nebyly zjištěny žádné pohyblivé buňky, je to označeno absencí černých čar.
Diskuse
Příbuzné rodu Shewanella tvoří heterotrofní fakultativní aeroby schopné využívat širokou škálu organických a anorganických látek jako zdroje energie. Mnoho kmenů rodu Shewanella bylo izolováno z vodního sloupce a sedimentů na lokalitách po celém světě. Kromě rozpustných akceptorů elektronů mohou mnohé z těchto organismů za anoxických podmínek respirovat široký sortiment přirozeně se vyskytujících nerozpustných oxidů kovů.
V naší studii shlukovacího chování u osmi druhů rodu Shewanella jsme pomocí sledování buněk pomocí počítačové analýzy a testu časové řady navázání buněk na MnO2 nebo Fe(OH)3 prokázali, že pět z osmi druhů rodu Shewanella je schopno tohoto chování CN32, ANA-3, W3-18-1, SB2B a PV-4) a že některé kmeny (W3-18-1, SB2B) vykazují za těchto podmínek preferenci jednoho oxidu kovu před druhým (obr. 4). Tato zjištění jsou v souladu s představou, že se tyto bakterie přizpůsobily převažujícímu nerozpustnému akceptoru elektronů, který se nachází v prostředí, z něhož byly izolovány. Navíc tři kmeny Shewanella, které byly izolovány z vodního sloupce, se v reakci na IEA nesdružovaly. Zda tyto rozdíly vyplývají z nepřítomnosti genů zapojených do EET (jako u OS217) nebo z jiných důvodů, bude předmětem budoucích studií.
V jiných studiích se ukázalo, že noční růst kmenů Shewanella porovnávající rychlost redukce různých oxidů kovů v porovnání hlava na hlavě, redukuje přednostně oxidy kovů z jejich environmentální niky (Bretschger et al., 2007). Zde uvedené výsledky jsou v souladu s hypotézou, že se u druhů rodu Shewanella vyvinulo shromažďovací a připojovací chování odpovídající prostředí, ze kterého byly izolovány (tabulka 3). Zdá se například, že W3-18-1 vykazuje výrazný sklon ke shlukování v přítomnosti minerálů Fe(OH)3 původem z pacifického kontinentálního šelfu a k navázání na ně (Harris et al., 2012), zatímco SB2B preferuje MnO2 (Venkateswaran et al., 1998) podobně jako v amazonském říčním sedimentu.
Srovnání genomů v tabulce 1 naznačuje, že přítomnost oktahemového cytochromu (SO_4142) může být důležitá pro plavání v reakci na relativně „nízké“ redoxní potenciály (100-300 mV oproti Ag/AgCl) Fe(OH)3 . Zatímco mtrF (SO_1780) může být potřebný pro reakci na relativně „vyšší“ redoxní potenciály MnO2 (mezi 400 a 600 mV oproti Ag/AgCl). Tuto hypotézu by pak bylo možné testovat pomocí delečních mutantů v MR-1. Tato práce doplňuje předchozí práci Harris et al. (2012) tím, že odhaluje, že geny podílející se na čistém efektu pohybového chování vůči minerálům IEA nebo elektrodám se kumulují v příslušných časových škálách 1-30 min (obr. 2) (Harris et al., 2012). Naše zjištění pomocí techniky konfokální mikroskopie CAAP prokázala, že kongruence vede k pomalé migraci buněk směrem k minerálům IEA s případným přichycením. Toto chování je tedy vhodně označováno jako shlukování, protože popisuje shromažďování nebo hromadění pohyblivých bakterií v okolí IEA. Zde jsme ukázali, že geny mcp cache a mtrBC/omcA jsou nezbytné pro akumulaci a fenotyp přichycení u MR-1, kromě toho, že jsou zodpovědné za charakteristické plavecké vzorce zvýšené rychlosti a chování typu run-and-reversal, které bylo identifikováno dříve (Harris et al., 2012).
Výsledky experimentů s mutanty ΔcymA, ΔmtrBC/omcA a Δmcp_cache u MR-1 ukazují inhibici akumulace, přichycení a shlukového chování v reakci na všechny IEA. Lze tedy předpokládat, že přítomnost homologních genů EET (cymA, mtrB, mtrC, omcA) a genu pro chemotaxi přijímající methyl (mcp_cache) určuje fenotypové odpovědi, které pozorujeme u jiných Shewanella spp (tab. 3). Genetické srovnání kmenů, které by mohly reagovat na „nižší“ redoxní potenciál IEA, naznačuje, že periferní oktahemové cytochromy vnější membrány (např. SO_4142) mohou hrát určitou roli v reakci na „nižší“ redoxní potenciál IEA. Proto tato pozorování o trvalém plavání kolem specifických minerálů odpovídala specifickým genotypům různých druhů.
Během mnoha experimentů s MFC (Kotloski a Gralnick, 2013) a s dávkovými kulturami redukujícími kovy nebyla měřena taxe energie zprostředkovaná mimobuněčným přenosem elektronů nebo shromažďovací schopnost (Kotloski a Gralnick, 2013). Při pokusech s redukcí oxidů kovů mohla 3D distribuce buněk a buněčná pohyblivost výrazně ovlivnit redukci kovů v důsledku rychlosti třepačky inkubátoru, rozměrů kultivační baňky nebo míchání. Kongregační chování přímo ovlivňuje přichycení buněk k IEA. Vztah mezi kongregací a tvorbou nanodrátovitých přívěsků není dosud znám (Pirbadian et al., 2015), ačkoli parametry motility a kongregace jsou v těchto studiích sledovány jen zřídka (Gorby et al., 2005; Reguera et al., 2005). Transformace těchto základních shromažďovacích genů (cymA, mtrB, mtrA, omcA, mcp_pas a mcp_cache) do jiných druhů bakterií s jedním polárním bičíkem může jednoho dne zlepšit bioremediační schopnosti tím, že bude schopna vyvolat uchycení bakterií a kolonizaci povrchů, které by jinak byly obtížné nebo nemožné.
Materiál a metody
Kultivace a kmeny
V této studii byly zkoumány MR-1 a několik delečních mutantů vzniklých z MR-1 (tabulka 2) (Beliaev a Saffarini, 1998; Myers a Myers, 2002). Kmeny byly inokulovány z glycerolových zásob uchovávaných při -80 °C na Luria-Bertaniho (LB) destičky a pěstovány přes noc při 30 °C. Poté byly jednotlivé kolonie vybrány a inokulovány do 5 ml definovaného minimálního média (M1) (Bretschger et al., 2007) doplněného 18 mM laktátem jako zdrojem energie (Bretschger et al., 2007) v 15 ml zkumavkách (VWR International LLC, Randor, Pennsylvania, USA) a inkubovány horizontálně v třepačce (180 ot./min.) po dobu 48 h při 30 °C. Optická hustota byla měřena pomocí spektrofotometru (Unico 1100RS spectrophotometer, Dayton, New Jersey, USA). Vzorky buněk byly odebrány při OD600 0,5 ± 0,2 (po ~48 h). Při plaveckých pokusech byly odebrány vzorky z pětimililitrových kultur, když buňky dosáhly OD 0,4, smíchány s oxidy manganu nebo železa a zavedeny do skleněné kapiláry (0,02 × 0,20 mm) (Vitrocom, Mountain Lakes, New Jersey, USA), která byla poté uzavřena pomocí vakuového tuku, jak bylo popsáno dříve (Harris et al., 2010).
Syntéza minerálů
Zásobní roztok Fe(OH)3 byl připraven podle protokolu Cornella a Schwertmanna a poté ověřen rentgenovou defrakcí (Schwertmann a Cornell, 2008). Tato příprava koloidního MnO2 začala rozpuštěním 8 g KMnO4 ve 200 ml při využití všech možných bezpečnostních opatření. Roztok byl průběžně míchán pomocí magnetické míchačky na vysoké teplotě a zahříván pod teplotu varu. Poté bylo přidáno 5 ml 10 M hydroxidu sodného k neutralizaci kyseliny vzniklé reakcí. V samostatné baňce bylo v 75 ml destilované vody rozpuštěno 15 g chloridu manganatého. Nakonec byl roztok pomalu smíchán s roztokem manganistanu (v chemické digestoři) po dobu 75 min. Po ochlazení byl roztok následně promyt odstředěním a pětkrát propláchnut deionizovanou vodou (DI) (18 Meg-Ohm cm). Konečná sraženina se nechala vyschnout pomocí vakuového filtru v čisté lavici a vysoušela se 36 h. Vzniklé minerály byly analyzovány pomocí rentgenové difrakce, aby se potvrdila produkce Fe(OH)3 a MnO2 (Bretschger et al., 2007; Salas et al., 2010).
Suspendované částice minerálů byly smíchány s kulturou o konečné koncentraci 300 mg/ml MnO2 nebo Fe(OH)3 . Buňky byly poté kapilárou ponořeny do obdélníkových kapilár (0,02 × 0,20 mm) (Vitrocom, Mountain Lakes, New Jersey, USA). Zkumavky byly uzavřeny silikonovým vakuovým tukem (Dow Corning, Midland, Milwaukie, USA) a pozorovány světelnou mikroskopií, fluorescenční mikroskopií a konfokální mikroskopií.
Časosběrné experimenty s akumulací buněk po fotobuňkách (CAAP)
Všechny fluorescenčně značené kmeny (GFP) byly transformovány, jak bylo popsáno dříve (McLean et al., 2008) a poté pěstovány aerobně na definovaném minimálním médiu s 25 μg/ml kanamycinu a 18 mM laktátu po dobu 48 h při 30 °C. Pětimililitrové kultury byly odebrány, když buňky dosáhly OD 0,4, smíchány s oxidy manganu nebo železa a zavedeny do skleněné kapiláry (0,02 × 0,20 mm) (Vitrocom, Mountain Lakes, New Jersey, USA), která byla poté uzavřena pomocí vakuového tuku, jak bylo popsáno dříve (Harris et al., 2010). GFP označené buňky WT MR-1, ΔcymA, Δmcp_cache a ΔmtrBC/omcA byly vyběleny pomocí nastavení maximální intenzity světla s 60x objektivy, inverzního konfokálního mikroskopu Leica TCS SPE (Wetzlar, Německo) po dobu 15 min. Aby bylo zajištěno, že došlo k vybělení, časosběrně se každou minutu zachytila oblast obrazovky (vybraná oblast 250 × 250 μm), dokud se původní buňky neobjevily tmavé a okolní buňky zůstaly jasně fluoreskující. Poté byly pořízeny snímky s použitím excitace 588 nm a emise 530 nm. Časosběrné video celého řezu zkumavky bylo pořízeno pomocí softwaru Leica Imaging a funkce „auto focus“ po dobu následujících 3 h. Buňky byly také pozorovány v režimu procházejícího světla, aby se ověřilo, že vybělené buňky jsou pohyblivé a neporušené. Samostatná negativní kontrola s GFP označeným Δmcp_cache byla snímána po dobu 3 h. U této negativní kontroly nebyly pozorovány žádné buňky hromadící se v tmavé zóně, ani odbarvené buňky neobnovily fluorescenci GFP. Reakce celé kapiláry (výška 30 μm) byla zachycena pomocí časosběrné fotografie a pro každý časový bod byl stanoven součet buněk ve všech 20 zásobnících v ose z (pomocí níže uvedené metody počítačové analýzy).
Mikroskopické zachycení pohybu buněk
Metody sledování a analýzy bakterií byly totožné s předchozími studiemi využitými pro obrázek 3 a zaznamenanými v tabulce 3 (Harris et al., 2012). Počítačové i manuální stopy plavání bakterií byly standardizovány pomocí pravítka (100 μm). Z každého experimentu byla zaznamenána celková plavecká aktivita v rámci videosnímku – ekvivalentní zornému poli o rozměrech 107 × 193 μm – a video bylo časově normalizováno, aby se získala rychlost plavání v μm/s. Pro každou plaveckou dráhu bakterií bylo měřeno několik parametrů, jako je celková uražená vzdálenost, doba dráhy od okamžiku, kdy se bakterie poprvé objeví a zmizí, počet couvnutí, vzdálenost mezi každým couvnutím a oxidem kovu a vzdálenost mezi oxidem kovu a začátkem dráhy bakterií.
Analýza plavání
Buňky byly sledovány ručně z videodat (30 snímků/s), snímek po snímku, a měřeny pomocí softwaru ImageJ pro analýzu obrazu. Vstupní data zahrnují rychlost plavání, počáteční polohu bakterií vzhledem k nejbližšímu povrchu IEA a poloha každé události zvratu bakterií byla identifikována a zaznamenána s ohledem na vzdálenost od nejbližšího povrchu IEA (Obrázek 3 a Tabulka 3). Pro stanovení plavecké aktivity po kontaktu s IEA byly plovoucí buňky pro analýzu rozděleny do dvou skupin: buňky, které plavaly do vzdálenosti 2 μm od částice, byly považovány za „kontaktní“ a ty, které neplavaly do vzdálenosti 2 μm od povrchu částice, byly považovány za „nekontaktní“. Kromě výše popsaných metod sledování rukou byla experimentální data následně počítačově roztříděna a analyzována dříve popsaným algoritmem, jehož výsledkem byly vypočtené údaje o plavání (Crocker a Grier, 1996; Harris et al., 2012; Harris, 2013). Pro vytvoření obrázků 3, 4 byla všechna experimentální data ze čtyř samostatných biologických replikátů zkombinována, spojena a zprůměrována do binů podle vzdálenosti od částice IEA. Vzhledem k tomu, že tato data byla sdružena do vzdáleností, chybové úsečky se překrývají; mezi kmeny nelze provést srovnání frekvence zvratu, v dané vzdálenosti, protože tento způsob vizualizace vylučuje identifikaci experimentů.
Příspěvky autorů
HH: navrhl a provedl experimenty; HH a IS-A: napsali a revidovali článek; ES, ME-N a JM: pomohli navrhnout experimenty; WT: pomohl provést analýzu dat; JM a KN: revidovali rukopis.
Prohlášení o střetu zájmů
Autoři prohlašují, že výzkum byl prováděn bez jakýchkoli komerčních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
Poděkování
Zvláštní poděkování patří Mandy J. Ward a Gijsu Kuenenovi za rady ohledně výzkumu a návrhu experimentu. Děkujeme Meaghan Sullivanové za ruční analýzy sledování. Děkujeme Cecile Jourlin-Castelli, Samanthě Reed, Jun Li a Davidu Culleymu za poskytnutí mutantů Δmcp_cache, ΔmtrBC/omcA a ΔcymA. Financování této práce bylo zajištěno z programu Air Force Office of Scientific Research Award. Výzkum IS-A byl podpořen stipendiem NASA Planetary Biology Internship.
Doplňkový materiál
Doplňkový materiál k tomuto článku naleznete online na adrese: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02568/full#supplementary-material
Alexandre, G., Greer-Phillips, S., and Zhulin, I. B. (2004). Ekologická úloha energetických taxisů v mikroorganismech. FEMS Microbiol. Rev. 28, 113-126. doi: 10.1016/j.femsre.2003.10.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Baraquet, C., Théraulaz, L., Iobbi-Nivol, C., Méjean, V., and Jourlin-Castelli, C. (2009). Neočekávané chemoreceptory zprostředkovávají energetickou taxaci k akceptorům elektronů u Shewanella oneidensis. Mol. Microbiol. 73, 278-290. doi: 10.1111/j.1365-2958.2009.06770.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Beliaev, A. S., and Saffarini, D. A. (1998). Shewanella putrefaciens mtrB kóduje protein vnější membrány potřebný pro redukci Fe(III) a Mn(IV). J. Bacteriol. 180, 6292-6297.
PubMed Abstract | Google Scholar
Bencharit, S., and Ward, M. J. (2005). Chemotaktické reakce na kovy a anaerobní akceptory elektronů u Shewanella oneidensis MR-1. J. Bacteriol. 187, 5049-5053. doi: 10.1128/JB.187.14.5049-5053.2005
PubMed Abstract | CrossRef Full Text | Google Scholar
Bond, D. R., and Lovley, D. R. (2003). Výroba elektrické energie bakterií Geobacter sulfurreducens připojenou k elektrodám. Appl. Environ. Microbiol. 69, 1548-1555. doi: 10.1128/AEM.69.3.1548-1555.2003
PubMed Abstract | CrossRef Full Text | Google Scholar
Bowman, J. P., McCammon, S. A., Nichols, D. S., Skerratt, J. H., Rea, S. M., Nichols, P. D., et al. (1997). Shewanella gelidimarina sp. nov. a Shewanella frigidimarina sp. nov., nové antarktické druhy se schopností produkovat kyselinu eikosapentaenovou (20:5ω3) a růst anaerobně pomocí disimilační redukce Fe(III). Int. J. Syst. Bacteriol. 47, 1040-1047. doi: 10.1099/00207713-47-4-1040
PubMed Abstract | CrossRef Full Text | Google Scholar
Bretschger, O. (2008). Schopnost přenosu elektronů a metabolické procesy rodu Shewanella s aplikacemi na optimalizaci mikrobiálních palivových článků. University of Southern California, ProQuest Dissertations Publishing, 3325185.
Google Scholar
Bretschger, O., Obraztsova, A., Sturm, C. A., Chang, I. S., Gorby, Y. A., Reed, S. B., et al. (2007). Produkce proudu a redukce oxidů kovů divokým typem a mutanty Shewanella oneidensis MR-1. Appl. Environ. Microbiol. 73:7003. doi: 10.1128/AEM.01087-07
PubMed Abstract | CrossRef Full Text | Google Scholar
Brettar, I., Christen, R., and Höfle, M. G. (2002). Shewanella denitrificans sp nov., silně denitrifikační bakterie izolovaná z oxicko-anoxického rozhraní Gotlandské hlubiny v centrální části Baltského moře. Int. J. Syst. Evol. Microbiol. 52, 2211-2217. doi: 10.1099/00207713-52-6-2211
PubMed Abstract | CrossRef Full Text | Google Scholar
Burdige, D. J. (1993). Biogeochemie redukce manganu a železa v mořských sedimentech. Earth Sci. Rev. 35, 249-284. doi: 10.1016/0012-8252(93)90040-E
CrossRef Full Text | Google Scholar
Champine, J. E., Underhill, B., Johnston, J. M., Lilly, W. W., and Goodwin, S. (2000). Přenos elektronů v disimilační železo redukující bakterii Geobacter metallireducens. Anaerobe 6, 187-196. doi: 10.1006/anae.2000.0333
CrossRef Full Text | Google Scholar
Childers, S. E., Ciufo, S., and Lovley, D. R. (2002). Geobacter metallireducens přistupuje k nerozpustnému oxidu Fe(III) pomocí chemotaxe. Nature 416, 767-769. doi: 10.1038/416767a
PubMed Abstract | CrossRef Full Text | Google Scholar
Coursolle, D., and Gralnick, J. A. (2010). Modularita respirační dráhy Mtr kmene Shewanella oneidensis MR-1. Mol. Microbiol. 77, 995-1008. doi: 10.1111/j.1365-2958.2010.07266.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Crocker, J. C., and Grier, D. G. (1996). Metody digitální videomikroskopie pro studium koloidů. J. Colloid Interface Sci. 179, 298-310. doi: 10.1006/jcis.1996.0217
CrossRef Full Text | Google Scholar
El-Naggar, M. Y., Wanger, G., Leung, K. M., Yuzvinsky, T. D., Southam, G., Yang, J., et al. (2010). Elektrický transport podél bakteriálních nanodrátků ze Shewanella oneidensis MR-1. Proc. Natl Acad. Sci. U.S.A. 107, 18127-18131. doi: 10.1073/pnas.1004880107
PubMed Abstract | CrossRef Full Text | Google Scholar
Fredrickson, J. K., Romine, M. F., Beliaev, A. S., Auchtung, J. M., Driscoll, M. E., Gardner, T. S., et al. (2008). Towards environmental systems biology of Shewanella. Nat. Rev. Microbiol. 6, 592-603. doi: 10.1038/nrmicro1947
PubMed Abstract | CrossRef Full Text | Google Scholar
Fredrickson, J. K., Zachara, J. M., Kennedy, D. W., Dong, H., Onstott, T. C., Hinman, N. W., et al. (1998). Biogenic iron mineralization accompanying the dissimilatory reduction of hydrous ferric oxide by a groundwater bacterium. Geochim. Cosmochim. Acta 62, 3239-3257. doi: 10.1016/S0016-7037(98)00243-9
CrossRef Full Text | Google Scholar
Gao, H., Obraztova, A., Stewart, N., Popa, R., Fredrickson, J. K., Tiedje, J. M., et al. (2006). Shewanella loihica sp. nov. izolovaná z mikrobiálních rohoží bohatých na železo v Tichém oceánu. Int. J. Syst. Evol. Microbiol. 56, 1911-1916. doi: 10.1099/ijs.0.64354-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Gorby, Y. A., Beveridge, T. J., and Wiley, W. R. (2005). „Composition, reactivity, and regulation of extracellular metal-reducing structures (nanowires) produced by dissimilatory metal reducing bacteria,“ in Annual NABIR PI Meeting (Warrenton, VA).
Google Scholar
Gorby, Y. A., Yanina, S., McLean, J. S., Rosso, K. M., Moyles, D., Dohnalkova, A., et al. (2006). Elektricky vodivé bakteriální nanodrátky produkované kmenem Shewanella oneidensis MR-1 a dalšími mikroorganismy. Proc. Natl. Acad. Sci. U.S.A. 103, 11358-11363. doi: 10.1073/pnas.0604517103
PubMed Abstract | CrossRef Full Text | Google Scholar
Gross, B. J., and El-Naggar, M. Y. (2015). Kombinovaná elektrochemická a optická trappingová platforma pro měření rychlosti dýchání jednotlivých buněk na rozhraní elektrod. Rev. Sci. Instrum. 86:064301. doi: 10.1063/1.4922853
PubMed Abstract | CrossRef Full Text | Google Scholar
Harris, H. W. (2013). Identifikace nového bakteriálního senzorového mechanismu: Bacterial Insoluble Electron Acceptor Sensing: Characterization of Bacterial Insoluble Electron Acceptor Sensing. University of Southern California.
Google Scholar
Harris, H. W., El-Naggar, M. Y., Bretschger, O., Ward, M. J., Romine, M. F., Obraztsova, A. Y., et al. (2010). Elektrokineze je mikrobiální chování, které vyžaduje extracelulární transport elektronů. Proc. Natl. Acad. Sci. U.S.A. 107, 326-331. doi: 10.1073/pnas.0907468107
PubMed Abstract | CrossRef Full Text | Google Scholar
Harris, H. W., Naggar, M. Y., and Nealson, K. H. (2012). Chemotaxické proteiny Shewanella oneidensis MR-1 a složky elektron-transportního řetězce nezbytné pro shlukování v blízkosti nerozpustných akceptorů elektronů. Biochem. Soc. Trans. 40, 1167-1177. doi: 10.1042/BST20120232
PubMed Abstract | CrossRef Full Text | Google Scholar
Hau, H. H., and Gralnick, J. A. (2007). Ekologie a biotechnologie rodu Shewanella. Annu. Rev. Microbiol. 61, 237-258. doi: 10.1146/annurev.micro.61.080706.093257
PubMed Abstract | CrossRef Full Text | Google Scholar
Hsu, L., Masuda, S. A., Nealson, K. H., and Pirbazari, M. (2012). Evaluation of microbial fuel cell Shewanella biocathodes for treatment of chromate contamination (Hodnocení mikrobiálních palivových článků s biokatodami Shewanella pro úpravu znečištění chromany). R. Soc. Chem. Adv. 2, 5844-5855. doi: 10.1039/c2ra20478a
CrossRef Full Text | Google Scholar
Kan, J., Hsu, L., Cheung, A. C. M., Pirbazari, M., and Nealson, K. H. (2011). Produkce proudu bakteriálními společenstvy v mikrobiálních palivových článcích obohacených z odpadních kalů s různými donory elektronů. Environ. Sci. Technol. 45, 1139-1146. doi: 10.1021/es102645v
PubMed Abstract | CrossRef Full Text | Google Scholar
Kato, S., Nakamura, R., Kai, F., Watanabe, K., and Hashimoto, K. (2010). Respirační interakce půdních bakterií s (polo)vodivými minerály oxidu železa. Environ. Microbiol. 12, 3114-3123. doi: 10.1111/j.1462-2920.2010.02284.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Kotloski, N. J., and Gralnick, J. A. (2013). Flavinové raketoplány dominují extracelulárnímu přenosu elektronů bakterií Shewanella oneidensis. MBio 4:e00553. doi: 10.1128/mBio.00553-12
PubMed Abstract | CrossRef Full Text | Google Scholar
Kracke, F., Vassilev, I., and Krömer, J. O. (2015). Mikrobiální transport elektronů a úspora energie – základ pro optimalizaci bioelektrochemických systémů. Front. Microbiol. 6:575. doi: 10.3389/fmicb.2015.00575
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, J., Go, A. C., Ward, M. J., and Ottemann, K. M. (2010). Chemická zkouška chemotaxe bakterií v zátce je náchylná k falešně pozitivním reakcím. BMC Res. Notes 3:77. doi: 10.1186/1756-0500-3-77
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, J., Romine, M. F., and Ward, M. J. (2007). Identifikace a analýza vysoce konzervovaného shluku genů pro chemotaxi u druhů rodu Shewanella. FEMS Microbiol. Lett. 273, 180-186. doi: 10.1111/j.1574-6968.2007.00810.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, R., Tiedje, J. M., Chiu, C., and Worden, R. M. (2012). Rozpustné elektronové raketoplány mohou zprostředkovávat energetické taxislužby směrem k nerozpustným elektronovým akceptorům. Environ. Sci. Technol. 46, 2813-2820. doi: 10.1021/es204302w
PubMed Abstract | CrossRef Full Text | Google Scholar
Lovley, D. R., Coates, J. D., Blunt-Harris, E. L., Phillips, E. J. P., and Woodward, J. C. (1996). Huminové látky jako akceptory elektronů pro mikrobiální respiraci. Nature 382, 445-448. doi: 10.1038/382445a0
CrossRef Full Text | Google Scholar
Lovley, D. R., Giovannoni, S. J., White, D. C., Champine, J. E., Phillips, E. J., Gorby, Y. A. a další (1993). Geobacter metallireducens gen. nov. sp. nov., mikroorganismus schopný spojit úplnou oxidaci organických sloučenin s redukcí železa a dalších kovů. Arch. Microbiol. 159, 336-344. doi: 10.1007/BF00290916
PubMed Abstract | CrossRef Full Text | Google Scholar
Lovley, D. R., Holmes, D. E., and Nevin, K. P. (2004). Disimilační redukce Fe(III) a Mn(IV). Adv. Microb. Physiol. 49, 219-286. doi: 10.1111/j.1462-2920.2004.00593.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Marsili, E., Baron, D. B., Shikhare, I. D., Coursolle, D., Gralnick, J. A., Bond, D. R., et al. (2008). Shewanella vylučuje flaviny, které zprostředkovávají extracelulární přenos elektronů. Proc. Natl. Acad. Sci. U.S.A. 105, 3968-3973. doi: 10.1073/pnas.0710525105
PubMed Abstract | CrossRef Full Text | Google Scholar
McLean, J. S., Majors, P. D., Reardon, C. L., Bilskis, C. L., Reed, S. B., Romine, M. F., et al. (2008). Zkoumání struktury a metabolismu v biofilmech Shewanella oneidensis MR-1. J. Microbiol. Methods 74, 47-56. doi: 10.1016/j.mimet.2008.02.015
PubMed Abstract | CrossRef Full Text | Google Scholar
Meyer, T. E., Tsapin, A. I., Vandenberghe, I., de Smet, L., Frishman, D., Nealson, K. H., et al. (2004). Identifikace 42 možných genů pro cytochrom C v genomu Shewanella oneidensis a charakterizace šesti rozpustných cytochromů. Omics 8, 57-77. doi: 10.1089/153623104773547499
PubMed Abstract | CrossRef Full Text | Google Scholar
Mitchell, A. C., Peterson, L., Reardon, C. L., Reed, S. B., Culley, D. E., Romine, M. R., et al. (2012). Úloha cytochromů typu c vnější membrány MtrC a OmcA v produkci, akumulaci a oddělování buněk Shewanella oneidensis MR-1 během respirace na hematitu. Geobiology 10, 355-370. doi: 10.1111/j.1472-4669.2012.00321.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Murray, A. E., Lies, D., Li, G., Nealson, K., Zhou, J., and Tiedje, J. M. (2001). DNA/DNA hybridizace na mikročipy odhaluje genově specifické rozdíly mezi blízce příbuznými mikrobiálními genomy. Proc. Natl. Acad. Sci. U.S.A. 98, 9853-9858. doi: 10.1073/pnas.171178898
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Myers, J. M. (1997). Cloning and sequence of cymA, a gene encoding a tetraheme cytochrome c required for reduction of iron(III), fumarate, and nitrate by Shewanella putrefaciens MR-1. J. Bacteriol. 179, 1143-1152. doi: 10.1128/jb.179.4.1143-1152.1997
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Myers, J. M. (2002). MtrB Is required for the proper incorporation of the cytochromes OmcA and OmcB into the outer membrane of Shewanella putrefaciens MR-1. Appl. Environ. Microbiol. 68, 5585-5594. doi: 10.1128/AEM.68.11.5585-5594.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Nealson, K. H. (1988a). Bakteriální redukce manganu a růst s oxidem manganičitým jako jediným akceptorem elektronů. Science 240, 1319-1321. doi: 10.1126/science.240.4857.1319
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Nealson, K. H. (1988b). Mikrobiální redukce oxidů manganu: Interakce s železem a sírou. Geochim. Cosmochim. Acta 52, 2727-2732. doi: 10.1016/0016-7037(88)90041-5
CrossRef Full Text | Google Scholar
Myers, J. M., and Myers, C. R. (2001). Role cytochromů OmcA a OmcB vnější membrány Shewanella putrefaciens MR-1 při redukci oxidu manganičitého. Appl. Environ. Microbiol. 67, 260-269. doi: 10.1128/AEM.67.1.260-269.2001
PubMed Abstract | CrossRef Full Text | Google Scholar
Nealson, K. H., Moser, D. P., and Saffarini, D. A. (1995). Anaerobní chemotaxe akceptorů elektronů u Shewanella putrefaciens. Appl. Environ. Microbiol. 61, 1551-1554.
PubMed Abstract | Google Scholar
Okamoto, A., Kalathil, S., Deng, X., Hashimoto, K., Nakamura, R., and Nealson, K. H. (2014). Flaviny vylučované buňkami a vázané na membránové cytochromy diktují reakce přenosu elektronů na povrchy s různým nábojem a pH. Sci. Rep. 4:5628. doi: 10.1038/srep05628
PubMed Abstract | CrossRef Full Text | Google Scholar
Pirbadian, S., Barchinger, S. E., Leung, K. M., Byun, H. S., Jangir, Y., Bouhenni, R. A., et al. (2015). Bakteriální nanodrátky Shewanella oneidensis MR-1 jsou vnější membránou a periplazmatickým rozšířením extracelulárních komponent transportu elektronů. Biophys. J. 108:368a. doi: 10.1016/j.bpj.2014.11.2016
CrossRef Full Text | Google Scholar
Reguera, G., McCarthy, K. D., Mehta, T., Nicoll, J. S., Tuominen, M. T., Lovely, D. R., et al. (2005). Extracelulární přenos elektronů prostřednictvím mikrobiálních nanodrátků. Nature 435, 1098-1101. doi: 10.1038/nature03661
PubMed Abstract | CrossRef Full Text | Google Scholar
Salas, E. C., Berelson, W. M., Hammond, D. E., Kampf, A. R., and Nealson, K. H. (2010). Vliv bakteriálního kmene na produkty disimilační redukce železa. Geochim. Cosmochim. Acta 74, 574-583. doi: 10.1016/j.gca.2009.10.039
PubMed Abstract | CrossRef Full Text | Google Scholar
Saltikov, C. W., Cifuentes, A., Venkateswaran, K., and Newman, D. K. (2003). Detoxikační systém ars je výhodný, ale není nutný pro respiraci As(V) geneticky ovlivnitelným kmenem druhu Shewanella ANA-3. Appl. Environ. Microbiol. 69, 2800-2809. doi: 10.1128/AEM.69.5.2800-2809.2003
CrossRef Full Text | Google Scholar
Schwalb, C., Chapman, S. K., and Reid, G. A. (2003). Tetrahemový cytochrom CymA je nezbytný pro anaerobní dýchání s dimethylsulfoxidem a dusitany u Shewanella oneidensis. Biochemistry 42, 9491-9497. doi: 10.1021/bi034456f
PubMed Abstract | CrossRef Full Text | Google Scholar
Schwertmann, U., and Cornell, R. M. (2008). Oxidy železa v laboratoři. Weinheim: John Wiley & Sons.
Google Scholar
Shi, L., Richardson, D. J., Wang, Z., Kerisit, S. N., Rosso, K. M., Zachara, J. M., et al. (2009). Role cytochromů vnějších membrán bakterií Shewanella a Geobacter v extracelulárním přenosu elektronů. Environ. Microbiol. Rep. 1, 220-227. doi: 10.1111/j.1758-2229.2009.00035.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Venkateswaran, K., Dollhopf, M. E., Aller, R., Stackebrandt, E., and Nealson, K. H. (1998). Shewanella amazonensis sp. nov., nový kovy redukující fakultativní anaerob z amazonských šelfových bahen. Int. J. Syst. Bacteriol. 48, 965-972.
PubMed Abstract | Google Scholar
Venkateswaran, K., Moser, D. P., Dollhopf, M. E., Lies, D. P., Saffarini, D. A., MacGregor, B. J., et al. (1999). Polyphasic taxonomy of the genus Shewanella and description of Shewanella oneidensis sp. nov. Int. J. Syst. Bacteriol. 49, 705-724. doi: 10.1099/00207713-49-2-705
PubMed Abstract | CrossRef Full Text | Google Scholar
Ziemke, F., Höfle, M. G., Lalucat, J., and Rossellö-Mora, R. (1998). Reklasifikace Shewanella putrefaciens Owenovy genomické skupiny II jako Shewanella baltica sp. nov. Int. J. Syst. Bacteriol. 48, 179-186. doi: 10.1099/00207713-48-1-179
PubMed Abstract | CrossRef Full Text | Google Scholar
.