FunctionEdit
ABA blev oprindeligt anset for at være involveret i abscission, og det er derfor, det har fået sit navn. Man ved nu, at dette kun er tilfældet i et lille antal planter. ABA-medieret signalering spiller også en vigtig rolle i planternes respons på miljøstress og plantepatogener. Plantegenerne for ABA-biosyntese og sekvensen af ABA-signaliseringsvejen er blevet opklaret. ABA produceres også af nogle plantepatogene svampe via en biosyntesevej, der er forskellig fra ABA-biosyntesen i planter.
I forbindelse med forberedelsen til vinteren produceres ABA i de terminale knopper. Dette bremser plantevæksten og styrer bladprimordier til at udvikle skæl for at beskytte de hvilende knopper i den kolde årstid. ABA hæmmer også celledelingen i det vaskulære cambium og tilpasser sig de kolde forhold om vinteren ved at suspendere primær og sekundær vækst.
Abscisinsyre produceres også i rødderne som reaktion på nedsat vandpotentiale i jorden (som er forbundet med tør jord) og andre situationer, hvor planten kan være under stress. ABA translokaliseres derefter til bladene, hvor det hurtigt ændrer det osmotiske potentiale i stomatalvagtcellerne, hvilket får dem til at krympe og lukker stomata. Den ABA-inducerede lukning af stomata reducerer transpirationen (fordampning af vand ud af stomataerne) og forhindrer dermed yderligere vandtab fra bladene i perioder med lav vandtilgængelighed. Der blev fundet en tæt lineær korrelation mellem ABA-indholdet i bladene og deres konduktans (stomatal modstand) på basis af bladareal.
Sædspiring hæmmes af ABA i antagonisme med gibberellin. ABA forhindrer også tab af frøets hviletid.
Der er identificeret flere ABA-mutante Arabidopsis thaliana-planter, som er tilgængelige fra Nottingham Arabidopsis Stock Centre – både planter med mangel på ABA-produktion og planter med ændret følsomhed over for ABA-aktivitet. Planter, der er overfølsomme eller ufølsomme over for ABA, udviser fænotyper med hensyn til frøhvile, spiring og stomatalregulering, og nogle mutanter udviser hæmmet vækst og brune/gule blade. Disse mutanter afspejler ABA’s betydning for frøspiring og tidlig embryonudvikling.
Pyrabactin (en pyridyl-holdig ABA-aktivator) er en naftalensulfonamid-hypokotylcelleudvidelseshæmmer, som er en agonist af ABA-signalvejen i frø. Det er den første agonist af ABA-signalvejen, som ikke er strukturelt beslægtet med ABA.
HomeostaseBearbejd
BiosynteseBearbejd
Abscisinsyre (ABA) er et isoprenoid plantehormon, som syntetiseres i den plastidale 2-C-methyl-D-erythritol-4-fosfat (MEP)-vej; I modsætning til de strukturelt beslægtede sesquiterpener, som dannes fra den af mevalonsyre afledte forløber farnesyldiphosphat (FDP), dannes ABA’s C15-rygsøjle efter spaltning af C40-karotenoider i MEP. Zeaxanthin er den første ABA-prækursor; en række enzymkatalyserede epoxideringer og isomeriseringer via violaxanthin og endelig spaltning af C40-karotenoidet ved en dioxygeneringsreaktion giver den proximale ABA-prækursor, xanthoxin, som derefter yderligere oxideres til ABA via abscisisk aldehyd.

Abamin er blevet designet, syntetiseret, udviklet og derefter patenteret som den første specifikke ABA-biosyntesehæmmer, hvilket gør det muligt at regulere de endogene niveauer af ABA.
Placering og timing af ABA-biosyntesenRediger
- Fremsættes under udtørring af de vegetative væv og når rødderne støder på jordkomprimering.
- Syntetiseres i grønne frugter i begyndelsen af vinterperioden
- Syntetiseres i modnende frø, der etablerer dvale
- Mobil i bladet og kan hurtigt translokaliseres fra rødderne til bladene af transpirationsstrømmen i xylem
- Produceret som reaktion på miljøstress, f.eks. varmestress, vandstress, saltstress
- Syntetiseres i alle plantedele, f.eks, rødder, blomster, blade og stængler
- ABA syntetiseres i næsten alle celler, der indeholder kloroplaster eller amyloplaster
InaktiveringRediger
ABA kan kataboliseres til fasesyre via CYP707A (en gruppe af P450-enzymer) eller inaktiveres ved glukosekonjugering (ABA-glukoseester) via enzymet AOG. Katabolisme via CYP707A’erne er meget vigtig for ABA-homeostase, og mutanter i disse gener akkumulerer generelt højere niveauer af ABA end linjer, der overudtrykker ABA-biosyntetiske gener. I jordbakterier er der rapporteret om en alternativ katabolisk vej, der fører til dehydrovomifoliol via enzymet vomifoliol dehydrogenase.
EffectsEdit
- Antitranspirant – Inducerer lukning af stomata, hvilket mindsker transpirationen for at forhindre vandtab.
- Hæmmer frugtmodning
- Er ansvarlig for frøets dvale ved at hæmme cellevækst – hæmmer frøspiringen
- Hæmmer syntesen af Kinetin-nukleotid
- Nedregulerer enzymer, der er nødvendige for fotosyntese.
- virker på endodermis for at forhindre vækst af rødder, når de udsættes for saltholdige forhold
- forsinker celledeling
- Dormancy inducer – Det bruges til at fremkalde hvile i frøene.
- bruges som anti – transpirant – I tørkefølsomme områder er vandstress et alvorligt problem i landbrugsproduktionen. Derfor foreslås det at sprøjte ABA, der forårsager delvis lukning af spalteåbninger i nogle få dage for at reducere transpirationstabet af vand
Signal kaskadeRediger
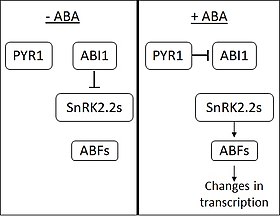
I fravær af ABA, phosphatasen ABI1-INSENSITIVE1 (ABI1) hæmmer virkningen af SNF1-relaterede proteinkinaser (underfamilie 2) (SnRK2s). ABA opfattes af membranproteinerne PYRABACTIN RESISTANCE 1 (PYR1) og PYR1-lignende membranproteiner. Ved ABA-binding binder PYR1 sig til og hæmmer ABI1. Når SnRK2 frigøres fra inhiberingen, aktiverer de flere transkriptionsfaktorer fra ABA RESPONSIVE ELEMENT-BINDING FACTOR-familien (ABF). ABF’erne forårsager derefter ændringer i ekspressionen af et stort antal gener. Man mener, at ca. 10 % af plantens gener reguleres af ABA.