For bedre at forstå denne mekanisme er der nedenfor en simpel opdeling af lugtvejen. Når mennesker tygger, skubbes flygtige smagsforbindelser gennem nasopharanx og lugtreceptorer.
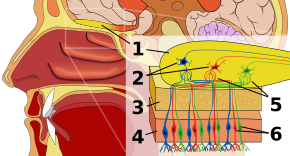
Olfaktorisk epitelRediger
Det første stop i det olfaktoriske system er det olfaktoriske epitel, eller væv, der hviler på taget af næsehulen, og som huser lugtreceptorer. Lugtreceptorer er bipolære neuroner, der binder lugtstoffer fra luften og samles ved lugtenerven, inden de sender axoner videre til dendriterne af mitralceller i lugtekolben. Sensoriske receptorer i mund og næse er polariseret i hviletilstand, og de depolariseres som reaktion på en ændring i omgivelserne, f.eks. når de kommer i kontakt med lugtmolekyler. Lugtmolekyler, der består af kulbrintekæder med funktionelle grupper, binder sig til sensoriske receptorer i næse og mund. De funktionelle gruppers egenskaber omfatter: (1) kulstofkædens længde, (2) terminalgruppe, som stemmer overens med de forskelle, der er forbundet med forskellige lugte, (3) sidegruppe, (4) kiralitet, (5) form og (6) størrelse. Når lugtmolekyler binder sig til sensoriske receptorer, gør de det i overensstemmelse med disse egenskaber. Hver lugtcelle har en enkelt type receptor, men denne receptor kan være “bredt indstillet”, og lugtmolekyler interagerer yderligere på receptorniveau, hvilket betyder, at et lugtmolekyle alene i visse tilfælde ikke binder sig til en receptor, men i tilstedeværelse af et andet lugtmolekyle vil det oprindelige molekyle binde sig og således kun skabe en lugtoplevelse i tilstedeværelse af det andet molekyle.
LugtbulbenRediger
I lugtbulben er lugtmolekylerne kortlagt rumligt. Disse rumlige repræsentationer er kendt som “lugtbilleder”. Den rumlige repræsentation muliggør lateral hæmning eller kontrastforstærkning og forstærkningskomprimering. Kontrastforstærkning er følsom over for ændringer og fremhæver stimuli i hjernen, der ændrer sig i stedet for at være i hvile. Gainkompression øger følsomheden over for stimuli med lav intensitet, mens følsomheden over for stimuli med høj intensitet mindskes. Lugtbulben, som forskerne stadig er i de indledende faser af deres forståelse af, adskiller lugtesansen fra andre sanser, fordi den markerer en afvigelse i sansevejen fra det, der er karakteristisk for alle andre sanser. Alle ikke-lugtsanselige sanseinformationer passerer nemlig gennem thalamus efter receptorniveauet, men det faktum, at lugtinformationer i stedet kommer ind i sit eget specialiserede område, kunne tyde på lugtens primitive historie og/eller en særskilt type behandling af lugtinformationer på deres vej til cortex. Lugtbulben rummer glomeruli, eller celleknudepunkter, hvor tusindvis af receptorer af samme type, ud over mitralceller, konvergerer. Denne organisation gør det muligt at repræsentere en stor mængde information kortfattet uden at kræve et lige så stort antal receptortyper. Den resulterende kombination af lugtinformationer kaldes et lugtbillede på niveauet af lugtekolben.
Billeddannelse i lugtekolbenRediger
2DG-metodeRediger
I 1977 udviklede biokemikeren Lou Sokoloff, Seymour Kety og Floyd E. Bloom en måde at kortlægge aktivitet i hjernen på ved at spore rottehjernens metabolisering af ilt. Nerveceller kræver ilt og glukose til energi. 2-deoxyglucose (2DG) er en radioaktiv glucoseisotop, som kan spores i hjernen, da den efterlader et spor i cellen, hvor den normalt ville blive omsat til energi, hvis det var glucose. Efter stimulering af et bestemt område af celler kan røntgenfotografier skives for at afsløre, hvilke celler der var aktive, især ved synaptiske knudepunkter.
Funktionel magnetisk resonansafbildning (fMRI) kan også bruges til at måle stofskiftet i en lugt. Denne metode er ikke terminal som 2-deoxyglukosemetoden, så et dyr kan måles med mange lugte, og de resulterende billeder kan sammenlignes.
Metoden med grønt fluorescerende proteinRediger
Endeligt kan man ved metoden med grønt fluorescerende protein genetisk manipulere mus til at udtrykke et protein i aktive neuroner, og et kamera kan derefter placeres inde i musens kranie for at måle aktiviteten.
ResultaterRediger
Disse metoder afslører især, at organiseringen af lugtinformation i løgbulben er rumlig. Lignende molekylære mønstre resulterer i lignende aktiveringsmønstre med hensyn til glomeruli, og glomeruli, der er tættere på hinanden, koder for lignende træk af lugtinformation.
Olfaktorisk cortexRediger
Den tre-lagede olfaktoriske cortex, der indeholder pyramidale celler, er det næste benchmark på lugtvejen. En pyramidecelle modtager information fra en mangfoldighed af mitralceller fra olfaktorisk pære, hvilket gør det tidligere organiserede glomerulære mønster fordelt i den olfaktoriske cortex. Denne spredning af mitralcelleinformation giver mulighed for selvudløsende feedbackforbindelser, lateral excitation og selv- og lateralinhibering. Disse processer bidrager til hebbisk læring, der er opkaldt efter Donald O. Hebb, og som ofte forenkles med ordsproget “neuroner, der skyder sammen, er forbundet med hinanden”. Langtidspotentiering, den neurale mekanisme for hebbisk læring, gør det muligt at danne hukommelse på pyramidecelleniveau. Hebbiansk læring er således hovedsagelig det fænomen, hvorved den olfaktoriske cortex “husker” resultatet af kombinationer af duftmolekyler og gør det muligt at genkende tidligere fornemmede kombinationer hurtigere end nye kombinationer ved at matche dem med lagrede input. De resulterende lugte, som tidligere blev kaldt lugtbilleder, lagres i olfaktorisk cortex med henblik på genkendelse og kaldes nu for lugtobjekter. Erfaring styrker derfor signal-støj-forholdet, idet et tidligere sanset lugtobjekt lettere kan skelnes fra en større baggrundsstøj.
Orbitofrontal cortexRediger
Den orbitofrontale cortex (OFC) er den endelige destination for lugtinformationerne og er der, hvor den bevidste lugtopfattelse opstår. Lugtinformationer kommer ind direkte efter at have passeret den olfaktoriske cortex, hvilket markerer forskellen fra andre sanseinformationer, der først passerer gennem thalamus. OFC er placeret dorsalt i forhold til den præfrontale cortex, hvilket giver lugtinformationer direkte input til den præfrontale cortex, eller det vigtigste beslutningsområde i hjernen. Der er kun tre typer neuroner, som lugtmolekyler passerer, inden de når frem til OFC: de olfaktoriske receptorceller i det olfaktoriske epitel, mitralceller og olfaktoriske pyramidale neuroner.
På OFC-niveau sker der associationer med andre hjerneområder, herunder input fra munden (somatosensation), følelsesmæssigt input (amygdala), visuel information og evaluerende information (præfrontal cortex). OFC er ansvarlig for selektiv lugtindstilling, sammensmeltning af sensoriske domæner og hedoniske vurderinger af lugte.