Para comprender mejor este mecanismo, a continuación se ofrece un sencillo desglose de la vía olfativa. Cuando los humanos mastican, los compuestos volátiles del sabor son empujados a través del nasofaringe y los receptores del olfato.
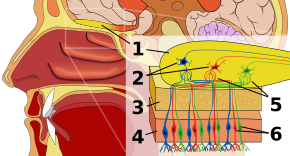
Epitelio olfativoEditar
La primera parada del sistema olfativo es el epitelio olfativo, o tejido que descansa en el techo de la cavidad nasal y que alberga los receptores del olor. Los receptores olfativos son neuronas bipolares que se unen a los odorantes del aire y se congregan en el nervio olfativo antes de pasar los axones a las dendritas de las células mitrales del bulbo olfativo. Los receptores sensoriales de la boca y la nariz están polarizados en estado de reposo y se despolarizan en respuesta a algún cambio en el entorno, como entrar en contacto con moléculas de olor. Las moléculas de olor, formadas por cadenas de hidrocarburos con grupos funcionales, se unen a los receptores sensoriales de la nariz y la boca. Las propiedades de los grupos funcionales incluyen: (1) la longitud de la cadena de carbono, (2) el grupo terminal, que concuerda con las diferencias asociadas a los distintos olores, (3) el grupo lateral, (4) la quiralidad, (5) la forma y (6) el tamaño. Cuando las moléculas olorosas se unen a los receptores sensoriales, lo hacen en función de estas propiedades. Cada célula olfativa tiene un único tipo de receptor, pero ese receptor puede estar «ampliamente sintonizado» y las moléculas de olor interactúan además a nivel del receptor, lo que significa que, en ciertos casos, una molécula de olor por sí sola puede no unirse a un receptor, pero en presencia de otra molécula de olor, la original se uniría y crearía así una sensación de olor sólo en presencia de la segunda molécula.
Bulbo olfativoEditar
En el bulbo olfativo, las moléculas de olor se representan espacialmente. Estas representaciones espaciales se conocen como «imágenes olfativas». La representación espacial permite la inhibición lateral, o el aumento del contraste y la compresión de la ganancia. El aumento del contraste es sensible al cambio y resalta los estímulos del cerebro que están cambiando y no en reposo. La compresión de la ganancia aumenta la sensibilidad a los estímulos de baja intensidad y disminuye la sensibilidad a los estímulos de alta intensidad. El bulbo olfativo, aunque todavía se encuentra en las primeras fases de su comprensión por parte de los investigadores, distingue al olfato de otros sentidos porque marca una desviación en la vía sensorial respecto a lo que es característico de todos los demás sentidos. A saber, toda la información sensorial no olfativa pasa por el tálamo después del nivel de los receptores, pero el hecho de que la información olfativa entre en cambio en su propia área especializada podría sugerir la historia primitiva del olfato y/o un tipo distinto de procesamiento de la información olfativa en su camino hacia la corteza. El bulbo olfativo alberga glomérulos, o uniones celulares, en los que confluyen miles de receptores del mismo tipo, además de células mitrales. Esta organización permite representar de forma concisa una gran cantidad de información sin requerir un número igualmente grande de tipos de receptores. La combinación resultante de la información de los olores se denomina imagen de los olores a nivel del bulbo olfativo.
Imágenes en el bulbo olfativoEditar
Método 2DGEditar
En 1977, el bioquímico Lou Sokoloff, Seymour Kety y Floyd E. Bloom desarrollaron una forma de cartografiar la actividad en el cerebro mediante el seguimiento de la metabolización del oxígeno en el cerebro de la rata. Las células nerviosas necesitan oxígeno y glucosa para obtener energía. La 2-deoxiglucosa (2DG) es un isótopo radiactivo de la glucosa que puede rastrearse en el cerebro, ya que deja un rastro en la célula donde normalmente se metabolizaría para obtener energía si fuera glucosa. Tras la estimulación de una determinada región de células, se pueden obtener fotografías de rayos X para revelar qué células estaban activas, sobre todo en las uniones sinápticas.
La resonancia magnética funcional (fMRI) también puede utilizarse para medir el metabolismo de un olor. Este método no es terminal como el método de la 2-deoxiglucosa, por lo que un animal puede ser medido con muchos olores, y las imágenes resultantes pueden ser comparadas.
Método de la proteína verde fluorescenteEditar
Por último, el método de la proteína verde fluorescente ingenieriza genéticamente a los ratones para que expresen una proteína en las neuronas activas, y entonces se puede colocar una cámara dentro del cráneo del ratón para medir la actividad.
ConclusionesEditar
Estos métodos revelan, sobre todo, que la organización de la información olfativa en el bulbo olfativo es espacial. Patrones moleculares similares dan lugar a patrones de activación similares con respecto a los glomérulos, y los glomérulos que están más juntos codifican características similares de la información olfativa.
Corteza olfativaEditar
La corteza olfativa de tres capas, que contiene células piramidales es el siguiente punto de referencia en la vía olfativa. Una célula piramidal recibe información de una multiplicidad de células mitrales del bulbo olfativo, haciendo que el patrón glomerular previamente organizado se distribuya en la corteza olfativa. Esta dispersión de la información de las células mitrales permite conexiones de retroalimentación autoexcitatorias, excitación lateral y autoinhibición lateral. Estos procesos contribuyen al aprendizaje Hebbiano, llamado así por Donald O. Hebb, y que suele simplificarse con el dicho «las neuronas que se disparan juntas se conectan». La potenciación a largo plazo, el mecanismo neuronal del aprendizaje hebbiano, permite la formación de la memoria a nivel de las células piramidales. El aprendizaje hebbiano es, por tanto, el fenómeno por el que el córtex olfativo «recuerda» la salida de las combinaciones de moléculas de olor y permite reconocer las combinaciones previamente percibidas más rápidamente que las nuevas, al hacerlas coincidir con la entrada almacenada. Los olores resultantes, que antes se denominaban imágenes de olor, se almacenan en la corteza olfativa para su reconocimiento y se denominan ahora objetos de olor. La experiencia, por tanto, refuerza la relación señal-ruido en el sentido de que un objeto oloroso previamente percibido puede distinguirse más fácilmente frente a un mayor ruido de fondo.
Corteza orbitofrontalEditar
La corteza orbitofrontal (OFC) es el destino final de la información olfativa y es donde surge la percepción consciente del olor. La información olfativa entra directamente después de pasar por la corteza olfativa, lo que marca la diferencia con otras informaciones sensoriales que pasan primero por el tálamo. El córtex olfativo está situado dorsalmente al córtex prefrontal, lo que permite que la información olfativa entre directamente en el córtex prefrontal, que es la principal área de toma de decisiones del cerebro. Sólo hay tres tipos de neuronas por las que pasan las moléculas olfativas antes de llegar a la OFC: las células receptoras olfativas del epitelio olfativo, las células mitrales y las neuronas piramidales olfativas.
A nivel de la OFC, se realizan asociaciones con otras áreas cerebrales, incluyendo la entrada de la boca (somatosensación), la entrada emocional (amígdala), la información visual y la información evaluativa (corteza prefrontal). La OFC es responsable de la sintonización selectiva de olores, la fusión de dominios sensoriales y las evaluaciones hedónicas de los olores.