FonctionEdit
On pensait à l’origine que l’ABA était impliqué dans l’abscission, d’où son nom. On sait maintenant que ce n’est le cas que chez un petit nombre de plantes. La signalisation médiée par l’ABA joue également un rôle important dans les réponses des plantes au stress environnemental et aux agents pathogènes des plantes. Les gènes végétaux de la biosynthèse de l’ABA et la séquence de la voie ont été élucidés. L’ABA est également produit par certains champignons phytopathogènes par une voie de biosynthèse différente de celle de l’ABA chez les plantes.
En préparation de l’hiver, l’ABA est produit dans les bourgeons terminaux. Cela ralentit la croissance des plantes et dirige les primordia de feuilles vers le développement d’écailles pour protéger les bourgeons dormants pendant la saison froide. L’ABA inhibe également la division des cellules dans le cambium vasculaire, s’ajustant aux conditions froides de l’hiver en suspendant la croissance primaire et secondaire.
L’acide abscissique est également produit dans les racines en réponse à la diminution du potentiel hydrique du sol (qui est associé à un sol sec) et à d’autres situations dans lesquelles la plante peut être stressée. L’ABA se transloque ensuite vers les feuilles, où il modifie rapidement le potentiel osmotique des cellules de garde stomatiques, provoquant leur rétrécissement et la fermeture des stomates. La fermeture des stomates induite par l’ABA réduit la transpiration (évaporation de l’eau par les stomates), empêchant ainsi les feuilles de perdre davantage d’eau lorsque la disponibilité en eau est faible. Une corrélation linéaire étroite a été trouvée entre la teneur en ABA des feuilles et leur conductance (résistance stomatique) sur une base de surface foliaire.
La germination des graines est inhibée par l’ABA en antagonisme avec la gibbérelline. L’ABA empêche également la perte de la dormance des graines.
Plusieurs plantes Arabidopsis thaliana mutantes de l’ABA ont été identifiées et sont disponibles auprès du Nottingham Arabidopsis Stock Centre – à la fois celles déficientes en production d’ABA et celles dont la sensibilité à son action est altérée. Les plantes hypersensibles ou insensibles à l’ABA présentent des phénotypes de dormance des graines, de germination, de régulation stomatique, et certains mutants présentent un retard de croissance et des feuilles brunes/jaunes. Ces mutants reflètent l’importance de l’ABA dans la germination des graines et le développement précoce de l’embryon.
La pyrabactine (un activateur de l’ABA contenant un pyridyle) est un inhibiteur de l’expansion cellulaire de l’hypocotyle à base de naphtalène sulfonamide, qui est un agoniste de la voie de signalisation de l’ABA des graines. C’est le premier agoniste de la voie ABA qui n’est pas structurellement apparenté à l’ABA.
HomeostasisEdit
BiosynthèseEdit
L’acide abscissique (ABA) est une hormone végétale isoprénoïde, qui est synthétisée dans la voie plastidiale du 2-C-méthyl-D-érythritol-4-phosphate (MEP) ; Contrairement aux sesquiterpènes structurellement apparentés, qui sont formés à partir du précurseur dérivé de l’acide mévalonique, le farnésyl diphosphate (FDP), le squelette C15 de l’ABA est formé après le clivage des caroténoïdes C40 dans la MEP. La zéaxanthine est le premier précurseur engagé de l’ABA ; une série d’époxydations et d’isomérisations catalysées par des enzymes via la violaxanthine, et le clivage final du caroténoïde en C40 par une réaction de dioxygénation donnent le précurseur proximal de l’ABA, la xanthoxine, qui est ensuite encore oxydée en ABA. via l’aldéhyde abscissique.

L’Abamine a été conçue, synthétisée, développée puis brevetée comme le premier inhibiteur spécifique de la biosynthèse de l’ABA, qui permet de réguler les niveaux endogènes d’ABA.
Localisation et moment de la biosynthèse de l’ABAModifié
- Libéré pendant la dessiccation des tissus végétatifs et lorsque les racines rencontrent une compaction du sol.
- Synthétisé dans les fruits verts au début de la période hivernale
- Synthétisé dans les graines en cours de maturation, établissant la dormance
- Mobile à l’intérieur de la feuille et peut être rapidement transloqué des racines aux feuilles par le courant de transpiration dans le xylème
- Produit en réponse au stress environnemental, tel que le stress thermique, le stress hydrique, le stress salin
- Synthétisé dans toutes les parties de la plante, par ex, racines, fleurs, feuilles et tiges
- L’ABA est synthétisé dans presque toutes les cellules qui contiennent des chloroplastes ou des amyloplastes
InactivationModification
L’ABA peut être catabolisé en acide phénique via le CYP707A (un groupe d’enzymes P450) ou inactivé par conjugaison au glucose (ABA-glucose ester) via l’enzyme AOG. Le catabolisme via les CYP707A est très important pour l’homéostasie de l’ABA, et les mutants de ces gènes accumulent généralement des niveaux plus élevés d’ABA que les lignées surexprimant les gènes de biosynthèse de l’ABA. Chez les bactéries du sol, une voie catabolique alternative conduisant au déhydrovomifoliol via l’enzyme vomifoliol déshydrogénase a été signalée.
EffetsEdit
- Antitranspirant – Induit la fermeture stomatique, diminuant la transpiration pour prévenir la perte d’eau.
- Inhibe le mûrissement des fruits
- Responsable de la dormance des graines en inhibant la croissance cellulaire – inhibe la germination des graines
- Inhibe la synthèse du nucléotide Kinetin
- Dérègle les enzymes nécessaires à la photosynthèse.
- Agit sur l’endoderme pour empêcher la croissance des racines lorsqu’elles sont exposées à des conditions salées
- Déclenche la division cellulaire
- Inducteur de dormance – Il est utilisé pour induire la dormance dans les graines .
- utilisé comme anti – transpirant – Dans les zones sujettes à la sécheresse , le stress hydrique est un problème sérieux dans la production agricole. Ainsi, des pulvérisations d’ABA sont suggérées qui provoquent une fermeture partielle des stomates pendant quelques jours , afin de réduire la perte transpirationnelle de l’eau
Cascade de signauxEdit
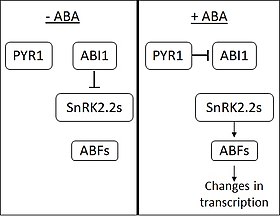
En l’absence d’ABA, la phosphatase ABI1-INSENSITIVE1 (ABI1) inhibe l’action des protéines kinases liées au SNF1 (sous-famille 2) (SnRK2s). L’ABA est perçu par les protéines membranaires PYRABACTIN RESISTANCE 1 (PYR1) et PYR1-like. Lors de la fixation de l’ABA, PYR1 se lie à ABI1 et l’inhibe. Lorsque les SnRK2 sont libérés de l’inhibition, ils activent plusieurs facteurs de transcription de la famille ABF (ABA RESPONSIVE ELEMENT-BINDING FACTOR). Les ABF provoquent ensuite des changements dans l’expression d’un grand nombre de gènes. On pense qu’environ 10% des gènes végétaux sont régulés par l’ABA.