Pour mieux comprendre ce mécanisme, une décomposition simple de la voie olfactive est fournie ci-dessous. Lorsque les humains mâchent, les composés d’arômes volatils sont poussés à travers le nasopharanx et les récepteurs d’odeur.
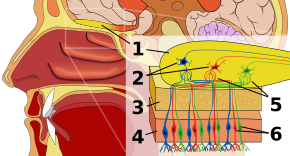
Épithélium olfactifModifié
Le premier arrêt du système olfactif est l’épithélium olfactif, ou tissu reposant sur le toit de la cavité nasale qui abrite les récepteurs olfactifs. Les récepteurs olfactifs sont des neurones bipolaires qui fixent les substances odorantes de l’air et se rassemblent au niveau du nerf olfactif avant de passer des axones aux dendrites des cellules mitrales du bulbe olfactif. Les récepteurs sensoriels de la bouche et du nez sont polarisés à l’état de repos, et ils se dépolarisent en réponse à un changement d’environnement, comme le contact avec des molécules odorantes. Les molécules odorantes, constituées de chaînes d’hydrocarbures avec des groupes fonctionnels, se lient aux récepteurs sensoriels du nez et de la bouche. Les propriétés des groupes fonctionnels comprennent (1) la longueur de la chaîne carbonée, (2) le groupe terminal, qui concorde avec les différences associées aux différentes odeurs, (3) le groupe latéral, (4) la chiralité, (5) la forme et (6) la taille. Lorsque les molécules odorantes se lient aux récepteurs sensoriels, elles le font en fonction de ces propriétés. Chaque cellule olfactive possède un seul type de récepteur, mais ce récepteur peut être « largement accordé » et les molécules odorantes interagissent en outre au niveau des récepteurs, ce qui signifie que, dans certains cas, une molécule odorante seule peut ne pas se lier à un récepteur, mais en présence d’une autre molécule odorante, l’originale se lierait et créerait ainsi une sensation d’odeur uniquement en présence de la seconde molécule.
Bulbe olfactifEdit
Dans le bulbe olfactif, les molécules odorantes sont cartographiées dans l’espace. Ces représentations spatiales sont connues sous le nom d' »images olfactives ». La représentation spatiale permet l’inhibition latérale, ou le renforcement du contraste et la compression du gain. L’amélioration du contraste est sensible au changement et met en évidence les stimuli du cerveau qui changent plutôt que ceux qui sont au repos. La compression du gain augmente la sensibilité aux stimuli de faible intensité tout en diminuant la sensibilité aux stimuli de forte intensité. Le bulbe olfactif, bien que les chercheurs n’en soient encore qu’aux premiers stades de sa compréhension, distingue l’odorat des autres sens car il marque une déviation dans la voie sensorielle par rapport à ce qui est caractéristique de tous les autres sens. En effet, toutes les informations sensorielles non olfactives passent par le thalamus après le niveau des récepteurs, mais le fait que les informations olfactives entrent dans leur propre zone spécialisée pourrait suggérer l’histoire primitive de l’odorat et/ou un type distinct de traitement des informations olfactives sur leur chemin vers le cortex. Le bulbe olfactif abrite des glomérules, ou jonctions cellulaires, sur lesquels convergent des milliers de récepteurs de même type, en plus des cellules mitrales. Cette organisation permet de représenter de manière concise une grande quantité d’informations sans nécessiter un nombre tout aussi important de types de récepteurs. La combinaison d’informations olfactives qui en résulte est surnommée une image olfactive au niveau du bulbe olfactif.
Imagerie dans le bulbe olfactifModification
Méthode 2DGModification
En 1977, le biochimiste Lou Sokoloff, Seymour Kety et Floyd E. Bloom ont mis au point un moyen de cartographier l’activité du cerveau en suivant la métabolisation de l’oxygène par le cerveau du rat. Les cellules nerveuses ont besoin d’oxygène et de glucose pour produire de l’énergie. Le 2-désoxyglucose (2DG) est un isotope radioactif du glucose qui peut être suivi dans le cerveau car il laisse une trace dans la cellule où il serait normalement métabolisé pour produire de l’énergie s’il s’agissait de glucose. Après stimulation d’une certaine région de cellules, des photographies aux rayons X peuvent être tranchées pour révéler quelles cellules étaient actives, en particulier aux jonctions synaptiques.
L’imagerie par résonance magnétique fonctionnelle (IRMf) peut également être utilisée pour mesurer le métabolisme d’une odeur. Cette méthode n’est pas terminale comme l’est la méthode du 2-désoxyglucose, ainsi un animal peut être mesuré avec de nombreuses odeurs, et les images résultantes peuvent être comparées.
Méthode de la protéine fluorescente verteModification
Enfin, la méthode de la protéine fluorescente verte modifie génétiquement les souris pour qu’elles expriment une protéine dans les neurones actifs, et une caméra peut ensuite être placée à l’intérieur du crâne de la souris pour mesurer l’activité.
ConstatationsModifier
Ces méthodes révèlent, notamment, que l’organisation de l’information olfactive dans le bulbe olfactif est spatiale. Des modèles moléculaires similaires entraînent des modèles d’activation similaires en ce qui concerne les glomérules, et les glomérules qui sont plus proches les uns des autres codent des caractéristiques similaires de l’information olfactive.
Cortex olfactifEdit
Le cortex olfactif à trois couches, contenant des cellules pyramidales est le prochain repère sur la voie de l’odorat. Une cellule pyramidale reçoit des informations d’une multiplicité de cellules mitrales provenant du bulbe olfactif, ce qui rend le motif glomérulaire précédemment organisé distribué dans le cortex olfactif. Cette dispersion des informations des cellules mitrales permet des connexions de rétroaction auto-excitatrices, une excitation latérale et une auto-inhibition et une inhibition latérale. Ces processus contribuent à l’apprentissage hébbien, nommé d’après Donald O. Hebb, et souvent simplifié par l’expression « les neurones qui tirent ensemble se connectent ensemble ». La potentialisation à long terme, le mécanisme neuronal de l’apprentissage de Hebb, permet la formation de la mémoire au niveau des cellules pyramidales. L’apprentissage hébbien est donc essentiellement le phénomène par lequel le cortex olfactif « se souvient » de la sortie des combinaisons de molécules odorantes et permet de reconnaître les combinaisons précédemment détectées plus rapidement que les nouvelles en les faisant correspondre aux données stockées. Les odeurs résultantes, qui étaient auparavant appelées images d’odeurs et qui sont stockées dans le cortex olfactif en vue de leur reconnaissance, sont désormais appelées objets odorants. L’expérience renforce donc le rapport signal/bruit en ce sens qu’un objet odorant précédemment perçu peut être plus facilement distingué par rapport à un bruit de fond plus important.
Cortex orbitofrontalEdit
Le cortex orbitofrontal (OFC) est la destination finale des informations olfactives et c’est là que la perception consciente des odeurs apparaît. Les informations olfactives entrent directement après être passées par le cortex olfactif, ce qui marque la distinction avec les autres informations sensorielles qui passent d’abord par le thalamus. Le CFO est situé en position dorsale par rapport au cortex préfrontal, ce qui permet aux informations olfactives d’entrer directement dans le cortex préfrontal, ou la principale zone de décision du cerveau. Il n’y a que trois types de neurones que les molécules d’odeur traversent avant d’arriver à l’OFC : les cellules réceptrices olfactives de l’épithélium olfactif, les cellules mitrales et les neurones pyramidaux olfactifs.
Au niveau de l’OFC, des associations avec d’autres zones cérébrales se font, notamment des entrées provenant de la bouche (somatosensation), des entrées émotionnelles (amygdale), des informations visuelles et des informations évaluatives (cortex préfrontal). L’OFC est responsable de l’accord sélectif des odeurs, de la fusion des domaines sensoriels et des évaluations hédoniques des odeurs.