- Introduction
- Tulokset
- Solujen kerääntyminen ja kiinnittyminen mineraaleihin vaatii kemotaksis- ja ekstrasellulaarisen elektroninkuljetuksen geenejä S. oneidensis MR-1:ssä
- Characterizing Congregation around IEA in Other Shewanella Strains
- Keskustelu
- Materiaalit ja menetelmät
- Viljely ja kannat
- Mineraalien synteesi
- Solujen kerääntyminen valonvalossa tapahtuvan valonvalkaisun jälkeen (CAAP) Time-Lapse-kokeet
- Solujen liikkeiden mikroskooppinen talteenotto
- Uintianalyysi
- Tekijän myötävaikutukset
- Kiinnostusristiriitoja koskeva lausunto
- Kiitokset
- Supplementary Material
Introduction
Kahdeksankymmentäluvun lopulla Shewanella oneidensis MR-1 (Myers ja Nealson, 1988a) ja myöhemmin useat Geobacter lajit (Lovley et al., 1993; Champine et al.., 2000) osoitettiin kykenevän siirtämään elektroneja liukenemattomiin elektroniakseptoreihin (IEA), kuten liukenemattomiin metallioksideihin ja/tai varattuihin elektrodeihin: prosessia kutsutaan solunulkoiseksi elektroninsiirroksi (EET) (Myers ja Nealson, 1988b; Venkateswaran et al., 1999; Bond ja Lovley, 2003). Tämä kyky on herättänyt huomattavaa kiinnostusta biogeokemiallisessa kierrossa, bioremediaatiossa, korroosiossa, nanomateriaalien käsittelyssä ja energiantuotannossa (Bretschger et al., 2007; Kan et al., 2011; Hsu et al., 2012). Vaikka useiden mikrobiryhmien tiedetään kykenevän EET:hen, merkittäviä mekanistisia tutkimuksia on tehty vain kahdella mallijärjestelmällä, Shewanella (Fredrickson et al., 2008; Shi et al., 2009) ja Geobacter (Lovley et al.,
Tähän mennessä yli 100 muuta Shewanella-kantaa on eristetty hyvin erilaisista elinympäristöistä, kuten avoimesta vesipatsaasta, hiekkakiviliuskeesta, meri- ja makean veden sedimenteistä, öljyputkista, öljysuolaliuoksesta ja jopa Antarktiksen jään päällä olevista leväyhteisöistä (Hau ja Gralnick, 2007). Yli 20 lajin genomit on sekvensoitu kokonaan (Fredrickson et al., 2008). Useiden näistä lajeista on osoitettu kykenevän EET:stä IEA:han, kuten S. oneidensis MR-1, Shewanella putrefaciens CN32, S. sp. ANA-3, S. sp. W3-18-1, Shewanella amazonensis SB2B, Shewanella frigidimarina NCIMB 400 ja Shewanella loihica PV-4 (Fredrickson et al., 1998; Venkateswaran et al., 1998; Gao et al., 2006; Bretschger, 2008). Muut Shewanella-suvun jäsenet, kuten Shewanella baltica OS217 ja Shewanella denitrificans OS155 (taulukko 1), eivät kykene EET:hen (Brettar et al., 2002). Kaikista näistä kannoista vain MR-1:n IEA:n vasteena tapahtuvaa kokoontumista on tutkittu.

TAULUKKO 1. Shewanella spp. ja niiden alkuperäisen elinympäristön geneettinen vertailu.
Shewanella-suvussa MR-1:n EET-mekanismia on luonnehdittu laajimmin. MR-1 käyttää useita lähestymistapoja liukenemattomien IEA-yhdisteiden pelkistämiseen: (1) suora EET solunulkoisten multihem-sytokromien kautta (Beliaev ja Saffarini, 1998; Myers ja Myers, 2001, 2002; Meyer et al., 2004; Mitchell et al., 2012; Kracke et al., 2015). (Kuva 1A); (2) välitetty EET käyttäen liukoisia elektronisukkuloita, jotka ovat sitoutuneet kalvosytokromeihin (Lovley ym., 1996; Marsili ym., 2008; Li ym., 2012; Kotloski ja Gralnick, 2013; Okamoto ym, 2014); (3) välitetty EET, jossa hyödynnetään johtavia ulkokalvon jatkeita, jotka sisältävät sytokromeja (Gorby et al., 2006; El-Naggar et al., 2010); ja (4) johtavia solunulkoisia matriiseja, jotka sisältävät johtavia ja puolijohtavia mineraaleja (Kato et al., 2010). Kannasta MR-1 on tunnistettu useita geenejä, joiden on osoitettu olevan välttämättömiä EET:n kannalta (kuva 1A ja taulukko 2), mukaan lukien tetraemi-sytokromi c cymA (SO_4591) ja mtrBC/omcA-yhdistelmä (SO_1776, SO_1778 ja SO_1779), jotka koodaavat useiden anaerobisten elektroniakseptoreiden, mukaan lukien metallioksidien, pelkistämiseen tarvittavaa dekaemi-sytokromi c:n komponenttia ja tetraemi-sytokromi c:n yhdistelmää varten (Myers ym., 2001, 2002; Schwalb et al., 2003). Koska kaikki nämä mekanismit ovat riippuvaisia solun läheisyydestä IEA:han EET:n saamiseksi, on tärkeää ymmärtää solun aistiminen ja netto-uintimuutto kohti IEA:ta. Seurakunnan, redox-taksonien tai energiataksonien osalta MR-1:llä on tehty monia tutkimuksia sen monipuolisen elektroniakseptorin hyödyntämisen vuoksi (Bencharit ja Ward, 2005; Baraquet et ai., 2009; Harris et ai., 2010). Energiataksi on termi, joka kattaa laajasti aerotaksian, fototaksian, redox-taksian, vaihtoehtoisiin elektroniakseptoreihin kohdistuvan taksian ja kemotaksian hapettuviin substraatteihin (Alexandre ym., 2004) (Alexandre ym., 2004).
KUVA 1. MR-1:n elektroniakseptit. (A) Ekstrasellulaarinen elektronin kuljetus (EET) Shewanella oneidensis MR-1:ssä rakentaa protonimoottorivoimaa (pmf). Anaerobisissa olosuhteissa Shewanella-suvun jäsenet voivat kuljettaa elektroneja sisäkalvolta, periplasmasta, ulkokalvolta ja elektrodilta sytokromien ja menakinonien (MQ) ketjun kautta, joka tunnetaan yhteisesti solunulkoisena elektronin kuljetuksena (EET). EET-sytokromien ilmentyminen voi vaihdella ympäristössä saatavilla olevan elektroniakseptorin mukaan. Rautaa sisältävien c-tyypin heemiryhmien lukumäärä on merkitty (suluissa). Elektroni virtaa elektroninluovuttajista, kuten laktaatista, pelkistäviin kinoneihin (Q), jotka samanaikaisesti siirtävät H+:a periplasmaan rakentaen pmf:ää ja siirtävät samalla elektroneja CymA:lle. Liukoisten elektroniakseptorien puuttuessa elektronit siirtyvät MtrCAB:n ulkokalvokompleksiin. Tämä MtrCAB-kompleksi voi luovuttaa elektroneja suoraan pääteelektroniakseptorille, joko mineraali- tai anodielektrodille, tai flaviinimolekyylien kautta. pmf ohjaa ATP:n tuotantoa ja polaaristen lippulaivojen pyörimistä. ”Itsetunnistavat” metyyliä hyväksyvät kemotaksiaproteiinit (MCP) ohjaavat lippulaivojen pyörimistä kemotaksisignaalinsiirtojärjestelmän (B) kautta ja voivat havaita H+ -pitoisuuden muutokset metallien pelkistymisen aikana. (B) Todennäköinen kemotaksis-signaalinsiirtoreitti Shewanellassa. Vastauksena stimulaatioon MCP:n rakenne siirtyy männän tavoin, jolloin CheA:n autofosforylaatio hidastuu tai pysähtyy. CheY ja CheB eivät siis fosforyloidu, ja tämä CheY-P:n puute mahdollistaa sujuvan uinnin. Tällä stimulaatiolla on myös toinen vaikutus – CheB on inaktiivinen ilman fosforylaatiota (se ei voi toimia metyyliesteraasina), ja tämä mahdollistaa sen, että CheR-proteiini (jatkuvasti aktiivinen metyylitransferaasi) voi kilpailla ja metyloida vapaasti MCP:n dimeerin metyyliä vastaanottavan alueen (HAMP-domeeni). Tämä MCP:n metylaatio lisää CheA:n histidiinijäännöksen autofosforylaationopeutta. Näin ollen signaalinsiirtojärjestelmä pystyy kontrolloimaan lippusolujen kääntötaajuutta lisääntyneiden tai vähentyneiden ärsykkeiden läsnä ollessa, mikä johtaa ”juokse-ja-käänny”-uintien sarjaan.

Taulukko 2. Lippusolujen kääntötaajuus. Tekstissä kuvatut MR-1:n geenit.
On hyvin dokumentoitu, että MR-1-solujen kertyminen vasteena liukoisiin elektroniakseptoreihin on eräänlainen energiataksi, joka on riippuvainen H+ -virrasta ja protonien liikkeellepanovoiman muodostumisesta (Baraquet ym., 2009) (kuva 1). Osa tähän vasteeseen kuuluu nopeampi uinti, kuten on nähty myös elektronisukkuloiden, kuten riboflaviinin tai antrakinoni-2,6-disulfonaatin (AQDS), kanssa (Bencharit ja Ward, 2005; Harris ym., 2010; Li ym., 2012). Sitä vastoin solujen kerääntyminen IEA: n ympärille, jota on kutsuttu kokoontumiseksi (Nealson et ai., 1995), liittyy sekä lisääntyneeseen uintinopeuteen kosketuksessa IEA: n kanssa (jota kutsutaan elektrokinesiaksi) että lisääntyneisiin uintikäänteisiin PMF: n vähentyessä. Uinnin kääntöliikkeet mahdollistavat useita ohimeneviä solun ja IEA:n kohtaamisia (kestävät 1-100 ms), ja nopeutta, jolla uivat solut siirtyvät irreversiibeliin kiinnittymiseen IEA:han kongregaation aikana, ei ole vielä kvantifioitu.
Mekanismi (mekanismit), jota (joita) solut käyttävät IEA:iden paikantamiseen, on (ovat) edelleen epäselvä(i)ä (Nealson et al., 1995; Bencharit ja Ward, 2005; Harris et al., 2010). Varhaisissa raporteissa ehdotettiin, että toinen tutkittu malliorganismi Geobacter kerääntyy IEA:iden ympärille aistimalla pelkistyneiden metalli-ionien gradientin (Childers et al., 2002), mutta pelkistyneet metalli-ionit eivät kuitenkaan liity MR-1:n käyttämään aistimismekanismiin (Bencharit ja Ward, 2005). Hiljattain kuvattiin bakteerikongregaatio vasteena paikoillaan oleviin elektrodeihin, mikä viittaa siihen, että vasteen laukaisevana tekijänä on pikemminkin redox-anturointi kuin metalli-ionien aistiminen (Harris et al., 2010). Elektrodeihin sovelletut positiiviset potentiaalit (200-600 mV vs. Ag/AgCl) aiheuttivat MR-1:n kokoontumisen samalla tavalla kuin metallioksidien kanssa havaittiin (Harris et al., 2012). MnO2:ta sisältävien mineraalien redox-potentiaalit vaihtelevat välillä 400-600 mV vs. Ag/AgCl, ja ne ottavat elektroneja vastaan helpommin kuin Fe(OH)3-mineraalit, joiden vastaava pollautettu potentiaali on 100-300 mV vs. Ag/AgCl (Burdige, 1993). Aiemmassa tutkimuksessa MR-1:n tyypillisen uinnin IEA:n ympärillä oletettiin olevan kahden itsetunnistavan kemotaksireseptorin, metyyliä hyväksyvien proteiinien (MCP), säätelemää, Ca+ -kanavan kemotaksireseptorin (Cache) domeenin (SO_2240) ja Per/Arnt/Sim (PAS) domeenin (SO_1385) avulla, ja kemotaksisignaalia välittävän proteiinikinaasin CheA-3 (SO_3207) avulla, jotta solu voi ottaa näytteen pinnan redox-potentiaalista tai elektronien vastaanottokyvystä (taulukko 2 ja kuva 1B) (Harris et al., )., 2012). Ennen tätä tutkimusta ei tunnettu geenejä, jotka ovat vastuussa solujen liikkuvuuteen perustuvasta kiinnittymisestä IEA:lle ja sen ympärille.
Tässä raportissa tutkimme eri Shewanella-kantojen kokoontumismekanismia seuraamalla uintitapoja ja solujen kiinnittymistä MnO2:een ja Fe(OH)3:een. MR-1:n WT:n ja useiden deletiomutanttien seulonnan lisäksi seulottiin myös muita Shewanella-lajeja (ks. taulukko 1). Monien kantojen osoitettiin kykenevän kerääntymään sekä MnO2:n että FeOH3:n ympärille, kun taas toiset reagoivat selektiivisesti MnO2:een (SB2B) tai Fe(OH)3:een (W3-18-1). Kun solujen kiinnittymistä mineraalipintoihin seurattiin, havaittiin vahva korrelaatio solun kyvyn kokoontua ja solujen kiinnittymisen IEA:n pintaan välillä. Sen jälkeen vertasimme näiden lajien genomeja löytääksemme kandidaattigeenejä, jotka osallistuvat kokoontumisuinti-, kerääntymis- ja solujen kiinnittymisfenotyyppeihin vasteena spesifisille IEA-pinnoille.
Tulokset
Solujen kerääntyminen ja kiinnittyminen mineraaleihin vaatii kemotaksis- ja ekstrasellulaarisen elektroninkuljetuksen geenejä S. oneidensis MR-1:ssä
Viimeisimmissä varhaistutkimuksissa, jotka koskivat MR-1:n energiantaksista, hyödynnettiin menetelmää, joka koostui parveilevan levyn määrityksistä (”swarm plate assays”) (Nealson et al., 1995; Baraquet et al., 2009; Li et al., 2012). Tässä tarjoamme syvällisemmän luonnehdinnan näistä vielä tuntemattomista energiataksien mekanismeista käyttämällä kvantitatiivista menetelmää, jossa seurataan yksittäisten solujen uintia nestemäisen väliaineen läpi vasteena IEA:lle ja solujen kertymiselle valonvalkaisun jälkeen (CAAP) solujen kiinnittymisnopeuden mittaamiseksi (Li et al., 2010). CAAP-menetelmässä käytetään konfokaalimikroskoopin UV-laserin säteilytystä, jolla GFP-merkityt solut tummuvat (sammutetaan fluoresenssi) peruuttamattomasti tietyn IEA:n päällä ja sen ympärillä. Kvantifioimalla fluoresoivia soluja niiden siirtyessä pimennetylle vyöhykkeelle voidaan kvantifioida kertymis- ja kiinnittymisnopeudet ja vertailla eri kantoja. Jotta solua voidaan pitää kiinnittyneenä, liikkuvan solun sijainti tarkistetaan analysoimalla 3D-kuva näkökentästä. Jos solu pysähtyy mineraalipinnalle, solua pidetään ”kiinnittyneenä”. Paikallaan olevien MR-1-solujen ja pintojen välisten sidosten lujuutta mitattiin optisilla pinseteillä erillisessä tutkimuksessa (Gross ja El-Naggar, 2015). Tämän menetelmän avulla voidaan erottaa toisistaan suunnattu solujen kiinnittyminen ja satunnainen sähköstaattinen kiinnittyminen.
Kuten kuvasta 2 nähdään, WT MR-1-solut kiinnittyivät mineraalipinnalle 30 minuutin kuluttua jonkin verran suurella lukumäärän poikkeamalla, 2 655 ± 1 352 solua/mm2. Tänä aikana solut osoittavat ”touch and go” -uintia, jolloin ne ottavat ohimenevän kosketuksen MnO2-mineraalipintaan. Kahden tunnin kuluttua mineraalipintaan kiinnittyneiden uusien solujen määrä kasvoi 4 300 ± 584 soluun/mm2 mineraalipintaa, kun taas lisää liikkuvia soluja kerääntyi edelleen (videot S1, S2 ja S17). Mutantit, joilla on kolmoisdeleetio keskeisissä EET-geeneissä mtrBC/omcA (SO_1776, SO_1778, SO_1779), yksittäinen deletio cymA:ssa (SO_4591) ja tärkeimmässä energiatakson kemoreseptorissa MCP Cache (SO_2240), on aiemmin yhdistetty uintikongregaatiofenotyyppiin (Harris ym., 2012). Deletoituneet mutantit (ΔcymA, Δmcp_cache ja ΔmtrBC/omcA) olivat kaikki liikkuvia ja kykenivät kääntämään uintisuuntaa, mutta eivät kyenneet kongregaatioon ja osoittivat vain vähän tai ei lainkaan kiinnittymistä MnO2:een kokeen aikana (kuva 2C ja kuvat S1A,B). WT:n kertyminen valkaistulle vyöhykkeelle t = 2 h:n kohdalla oli keskimäärin 4300 ± 584 solua/mm2, kun taas Δmcp_cache- ja ΔmtrBC/omcA-mutanttien kertyminen oli vähäistä (Kuva 2C ja S1CD).
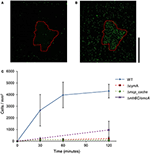
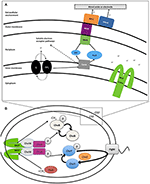
KUVA 2. WT:n kertyminen valkaistulle vyöhykkeelle t = 2 h. Uiva MR-1 voi vaeltaa kohti liukenemattomia elektroniakseptorimineraaleja ja kiinnittyä. Edustava konfokaalinen fluoresenssimikroskopiakuva WT MR-1 -soluista t = 0 ja t = 120 min (A,B). Uivia WT MR-1 GFP-soluja tuotiin MnO2-hiukkaseen (punainen katkoviiva) anaerobisessa suljetussa kapillaarissa. Hetkellä t = 0 kaikki solut valkaistuivat irreversiibelisti 250 x 250 μm:n alueella hiukkasen ympärillä (A). Tämän jälkeen laskettiin valkaistun alueen ulkopuolelta peräisin olevat fluoresoivat solut, jotka uivat kehykseen ja kiinnittyivät mineraalin pintaan. Oikealla oleva musta pystysuora asteikkopalkki edustaa 100 μm. Kaaviossa (C) verrataan WT MR-1:n kiinnittymistä MnO2-hiukkaseen 120 minuutin aikana kemotaksisuuteen ja solunulkoisen elektroninsiirron deleetioihin perustuviin mutaatioihin (MR-1 Δmcp_cache, ΔcymA ja ΔmtrBC/omcA). Virhepalkit sisältävät 2 std-poikkeamaa.
Characterizing Congregation around IEA in Other Shewanella Strains
Yhdeksän Shewanella-kannan kykyä kokoontua MnO2- ja Fe(OH)3-mineraalipintojen ympärille testattiin aikasarjamittauksella solujen kiinnittymisestä mineraaliin (taulukko 3 ja videot S3-S14). Kuten MR-1:n kohdalla, kaikilla yhdeksällä tutkituilla kannoilla oli yksi polaarinen lippulaite, ja uintisuunnan kääntäminen onnistui kääntämällä lippulaitteen kiertosuunta (tietoja ei ole esitetty). Saman kokeen uintiradat lajiteltiin kahteen erilliseen ryhmään uintiradan perusteella (taulukko 3): ne, jotka koskettivat liukenemattoman metallioksidin pintaa (uivat 2 μm:n sisällä) verrattuna niihin, jotka eivät koskettaneet (uivat >2 μm). Kosketukseen joutuneet uimarit, jotka osoittivat merkittävää lisäystä peruutustiheydessä ja uintinopeudessa verrattuna ei-kosketukseen joutuneeseen ryhmään (P < 0,05), on luokiteltu seurakuntakäyttäytymisen kannalta positiivisiksi, kuten on merkitty ylempänä olevilla kirjaimilla (taulukko 3).

TAULUKKO 3. Bakteerien uintinopeus ja kääntymisfrekvenssi metallioksidimineraalien ympärillä.
Kannat ANA3, CN32 ja PV-4 sekä MR-1 olivat MR-1:n lisäksi positiivisia kongregaatiokäyttäytymisen suhteen sekä MnO2:n että Fe(OH)3:n ympärillä (taulukko 3, kuva 3; videot S1-S8). Niiden kääntymisnopeus oli suurimmillaan, kun ne sijaitsivat 5-40 μm:n päässä MnO2- tai FeOH3-hiukkasen pinnasta (kuva 3). Kolme kantaa: OS217, NCIMB400 ja OS155 eivät olleet liikkuvia ajankohtana t = 30 min – vasteena Fe(OH)3- tai MnO2-mineraaleille (taulukko 3). Esimerkiksi MnO2:n kanssa kosketuksiin joutuneiden uivien MR-1-solujen kääntymistaajuus oli 0,94 ± 0,53 kääntymistä/s, kun taas kosketuksiin joutumattomien solujen kääntymistaajuus oli 0,62 ± 0,73 kääntymistä/s. Nopeus kasvoi myös MR-1:n kontaktiryhmässä, kun se oli 19,26 ± 11,2 μm/s ei-kontaktiryhmässä ja 24,37 ± 6 μm/s kontaktiryhmässä. Koska uintinopeus sekä lisääntyi tilastollisesti merkitsevästi että kääntyi niin, että solut pysyivät lähellä metallioksidipartikkelia, tämän kannan sanottiin olevan kongregaatiopositiivinen.
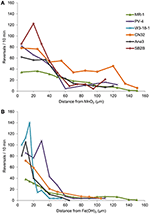
Kuva 3. (A,B) Uivien Shewanella-solujen kääntymistaajuus (y-akseli) vs. etäisyys (x-akseli) IEA-hiukkasesta. Yksittäisten seurattujen uintisolujen keskimääräiset kääntymistaajuudet on jaettu 5 μm:n väleihin x-akselia pitkin MnO2- (A) tai Fe(OH)3 -hiukkasella (B) tehdyissä kokeissa.
Shewanella spp:n uintiradat (30 s), jotka osoittivat mieltymystä metallioksidimineraaleihin, on korostettu kuvissa 4A-F. Kuva SB2B-solut eivät osoittaneet uintivasteensa Fe(OH)3:lle (kuva 4D), kun taas ne osoittivat aktiivista kokoontumista MnO2-hiukkasten ympärillä (kuva 4A). Sitä vastoin W3-18-1-solut kerääntyivät Fe(OH)3 -hiukkasten ympärille (kuva 4C), mutta osoittivat huomattavasti vähentynyttä uinti- ja kääntöaktiivisuutta MnO2:n ympärillä (kuva 4F; videot S9, S14).
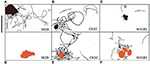
Kuva 4. (A-F) Paneeli kolmesta Shewanella spp. uintiradasta (30 s) osoitti mieltymystä metallioksidimineraaleihin. Tutkittiin kolmen liikkuvan Shewanella spp. uintikäyttäytymistä vasteena MnO2:lle (ylärivi-musta värillinen hiukkanen) ja Fe(OH)3:lle (alarivi-oranssi värillinen hiukkanen). Kongregaatio tapahtuu anaerobisissa olosuhteissa, uintiradat on esitetty mustalla. SB2B (vasemmalla), CN32 (keskellä) ja W3181 (oikealla). Solujen uintia seurattiin käsin. Mittakaavapalkki vasemmalla ylhäällä = 50 μm. Ruskea värimuoto edustaa MnO2:ta ja oranssi Fe(OH)3:a. Kun liikkuvia soluja ei ole havaittu, tämä näkyy mustien viivojen puuttumisena.
Keskustelu
Shewanella-suvun jäsenet koostuvat heterotrofisista, fakultatiivisista aerobeista, jotka kykenevät hyödyntämään energialähteenä monenlaisia orgaanisia ja epäorgaanisia aineita. Monia Shewanella-kantoja on eristetty vesipatsaan ja sedimentin elinympäristöistä eri puolilta maailmaa. Liukoisten elektroniakseptorien lisäksi monet näistä organismeista voivat hengittää anoksisissa olosuhteissa monenlaisia luonnossa esiintyviä liukenemattomia metallioksideja.
Tutkimuksessamme kahdeksan Shewanella-lajin kongregaatiokäyttäytymisestä käytimme solunseurantaa tietokoneanalyysillä ja solujen kiinnittymisen MnO2:een tai Fe(OH)3:een aikasarjamäärityksellä osoittaaksemme, että viisi kahdeksasta Shewanellasta kykeni tähän käyttäytymiseen (CN32, ANA-3, W3-18-1, SB2B ja PV-4) ja että jotkin kannat (W3-18-1, SB2B) osoittivat mieltymystä yhdelle metallioksidille toista enemmän näissä olosuhteissa (kuva 4). Tällaiset havainnot ovat sopusoinnussa sen kanssa, että nämä bakteerit ovat sopeutuneet vallitsevaan liukenemattomaan elektroniakseptoriin, jota esiintyy elinympäristössä, josta ne on eristetty. Lisäksi kolme Shewanella-kantaa, jotka eristettiin vesipatsaasta, eivät kokoontuneet vasteena IEA:lle. Se, johtuvatko nämä erot EET:hen osallistuvien geenien puuttumisesta (kuten OS217:llä) vai muista syistä, on tulevien tutkimusten aihe.
Muissa tutkimuksissa, joissa Shewanella-kantojen yön yli tapahtuvassa kasvussa verrattiin eri metallioksidien pelkistymisnopeuksia päätä päähän -vertailussa, osoitettiin, että ne pelkistävät metallioksidia mieluiten ympäristönsä kapeikosta (Bretschger et al., 2007). Tässä esitetyt tulokset ovat sopusoinnussa sen hypoteesin kanssa, että Shewanella-lajit ovat kehittäneet kokoontumis- ja kiinnittymiskäyttäytymisen, joka on yhdenmukainen niiden ympäristöjen kanssa, joista ne on eristetty (taulukko 3). Esimerkiksi W3-18-1 näyttäisi osoittavan merkittävää taipumusta kokoontumiseen Tyynenmeren mannerjalustalle tyypillisten Fe(OH)3-mineraalien läsnäollessa ja kiinnittymiseen niihin (Harris et al., 2012), kun taas SB2B suosii MnO2:ta (Venkateswaran et al., 1998), joka on samankaltainen kuin Amazonin jokisedimentissä.
Taulukossa 1 esitetty genomivertailu viittaa siihen, että oktahemisytokromin (SO_4142) läsnäolo voi olla tärkeää uimiselle vastauksena Fe(OH)3:n suhteellisen ”mataliin” redox-potentiaaleihin (100-300 mV vs. Ag/AgCl). Kun taas mtrF:ää (SO_1780) saatetaan tarvita MnO2:n suhteellisen ”korkeisiin” redox-potentiaaleihin (400-600 mV vs. Ag/AgCl) reagoimiseksi. Tätä hypoteesia voitaisiin sitten testata MR-1:n deletointimutaatioilla. Tämä työ täydentää Harrisin et al. (2012) aiempaa työtä paljastamalla, että geenit, jotka osallistuvat liikkuvuuskäyttäytymisen nettovaikutukseen kohti IEA-mineraaleja tai elektrodeja, ovat kumulatiivisia asiaankuuluvilla aikaskaaloilla 1-30 min (kuva 2) (Harris et al., 2012). Havaintomme CAAP-konfokaalimikroskopiatekniikalla osoittivat, että seurakunta johtaa solujen hitaaseen migraatioon kohti IEA-mineraaleja ja lopulta kiinnittymiseen. Näin ollen tätä käyttäytymistä kutsutaan sopivasti kongregaatioksi, koska se kuvaa liikkuvien bakteerien kokoontumista tai kerääntymistä IEA:n ympärille. Osoitimme tässä, että geenit mcp cache ja mtrBC/omcA ovat välttämättömiä kerääntymis- ja kiinnittymisfenotyypille MR-1:ssä sen lisäksi, että ne ovat vastuussa tyypillisistä uintikuvioista, jotka koostuvat lisääntyneestä nopeudesta ja juoksu- ja peruutustyyppisestä käyttäytymisestä, joka tunnistettiin aiemmin (Harris et al., 2012).
Kokeiden tulokset MR-1:n ΔcymA-, ΔmtrBC/omcA- ja Δmcp_cache-mutanteilla osoittavat, että kertyminen, kiinnittyminen ja kokoontumiskäyttäytyminen estyvät vasteena kaikille IEA:ille. Näin ollen voidaan olettaa, että homologisten EET-geenien (cymA, mtrB, mtrC, omcA) ja metyyliä hyväksyvän kemotaksigeenin (mcp_cache) läsnäolo määrittää fenotyyppiset vasteet, joita näemme muissa Shewanella spp:ssä (taulukko 3). Niiden kantojen geneettinen vertailu, jotka voivat reagoida ”alhaisemman” redox-potentiaalin IEA:han, osoittaa, että ulkokalvon perifeerisillä oktahemisytokromeilla (kuten SO_4142) voi olla jonkinlainen rooli ”alhaisemman” redox-potentiaalin IEA:han reagoimisessa. Näin ollen nämä havainnot jatkuvasta uimisesta tiettyjen mineraalien ympärillä vastasivat eri lajien erityisiä genotyyppejä.
Monien MFC- (Kotloski ja Gralnick, 2013) ja metallia pelkistävien panosviljelykokeiden aikana ei mitattu solunulkoisen elektroninsiirron välittämää energiantaksia eli kokoontumiskykyä (Kotloski ja Gralnick, 2013). Metallioksidipelkistyskokeissa solujen 3D-jakautuminen ja solujen liikkuvuus voivat vaikuttaa suuresti metallien pelkistymiseen inkubaattorin ravistimen nopeuden, viljelypullon koon tai sekoittumisen vuoksi. Kongregaatiokäyttäytyminen vaikuttaa suoraan solujen kiinnittymiseen IEA:iin. Kongregaation ja nanolankamaisen appendanssin muodostumisen välistä suhdetta ei vielä tunneta (Pirbadian et al., 2015), vaikka liikkuvuus- ja kongregaatioparametreja seurataan näissä tutkimuksissa harvoin (Gorby et al., 2005; Reguera et al., 2005). Näiden keskeisten kokoontumisgeenien (cymA, mtrB, mtrA, omcA, mcp_pas ja mcp_cache) muuntaminen muihin bakteerilajeihin, joilla on vain yksi polaarinen lippulehti, saattaa jonain päivänä parantaa bioremediaatiovalmiuksia, koska sillä voidaan indusoida bakteerien kiinnittyminen ja kolonisoituminen pinnoille, jotka muutoin olisivat vaikeita tai mahdotonta.
Materiaalit ja menetelmät
Viljely ja kannat
Tässä tutkimuksessa tutkittiin MR-1:tä ja useita MR-1:stä peräisin olevia deletoituneita mutantteja (taulukko 2) (Beliaev ja Saffarini, 1998; Myers ja Myers, 2002). Kannat inokuloitiin -80 °C:ssa säilytetyistä glyserolivarastoista Luria-Bertani (LB) -levyille ja kasvatettiin yön yli 30 °C:ssa. Yksittäiset pesäkkeet valittiin ja inokuloitiin 5 ml:aan määriteltyä minimaalista mediaa (M1) (Bretschger et al., 2007), jota täydennettiin 18 mM laktaatilla energianlähteenä (Bretschger et al., 2007) 15 ml:n putkissa (VWR International LLC, Randor, Pennsylvania, USA), ja niitä inkuboitiin vaakatasossa ravistimessa (180 rpm) 48 tuntia 30 °C:ssa. Optinen tiheys mitattiin spektrofotometrillä (Unico 1100RS spectrophotometer, Dayton, New Jersey, USA). Soluista otettiin näyte OD600:n ollessa 0,5 ± 0,2 (~48 tunnin kuluttua). Uintikokeissa näytteet otettiin viiden millilitran viljelmistä, kun solujen OD oli 0,4, sekoitettiin mangaani- tai rautaoksidien kanssa ja vietiin lasikapillaariin (0,02 × 0,20 mm) (Vitrocom, Mountain Lakes, New Jersey, USA), joka suljettiin tyhjiörasvalla aiemmin kuvatulla tavalla (Harris et al., 2010).
Mineraalien synteesi
Fe(OH)3-kantaliuos valmistettiin Cornellin ja Schwertmannin laatiman protokollan mukaisesti, minkä jälkeen se tarkistettiin röntgendiffraktiolla (Schwertmann ja Cornell, 2008). Tämä kolloidisen MnO2:n valmistus aloitettiin 8 g KMnO4:lla, joka oli liuotettu 200 ml:aan, samalla kun hyödynnettiin kaikkia mahdollisia varotoimenpiteitä. Liuosta sekoitettiin jatkuvasti magneettisella sekoituspuikolla korkealla ja kuumennettiin kiehumislämpötilan alapuolelle. Sitten lisättiin 5 ml 10 M natriumhydroksidia neutraloimaan reaktiossa syntynyt happo. Erillisessä pullossa 15 g mangaanikloridia liuotettiin 75 ml:aan tislattua vettä. Lopuksi liuosta sekoitettiin hitaasti permanganaattiliuokseen (kemiallisessa huuruhuoneessa) 75 minuutin ajan. Kun liuos oli jäähdytetty, se pestiin sentrifugoimalla ja huuhdeltiin deionisoidulla vedellä (DI) (18 Meg-Ohm cm) viisi kertaa. Lopullisen saostuman annettiin kuivua tyhjiösuodattimella puhtaassa penkissä ja kuivattiin 36 h. Syntyneet mineraalit analysoitiin röntgendiffraktiolla Fe(OH)3:n ja MnO2:n tuotannon vahvistamiseksi (Bretschger et al., 2007; Salas et al., 2010).
Suspendoituja mineraalihiukkasia sekoitettiin viljelmään siten, että loppupitoisuudeksi saatiin 300 mg/ML:aa MnO2:ta tai Fe(OH)3:a. Solut upotettiin sitten kapillaarisesti suorakulmaisiin kapillaariputkiin (0,02 × 0,20 mm) (Vitrocom, Mountain Lakes, New Jersey, Yhdysvallat). Putket suljettiin Silicon-tyhjiörasvalla (Dow Corning, Midland, Milwaukie, USA) ja niitä havainnoitiin valomikroskoopilla, fluoresenssimikroskoopilla ja konfokaalimikroskoopilla.
Solujen kerääntyminen valonvalossa tapahtuvan valonvalkaisun jälkeen (CAAP) Time-Lapse-kokeet
Kaikkia fluoresenssilla leimattuja kantoja (GFP) oli muunnettu aiemmin kuvattujen menetelmien mukaisesti (McLean et al., 2008) ja kasvatettiin sitten aerobisesti määritellyssä minimaalisessa väliaineessa, jossa oli 25 μg/ml kanamysiiniä ja 18 mM laktaattia, 48 tunnin ajan 30 °C:ssa. Viiden millilitran viljelmistä otettiin näyte, kun solujen OD oli 0,4, ne sekoitettiin mangaani- tai rautaoksidien kanssa ja vietiin lasikapillaariin (0,02 × 0,20 mm) (Vitrocom, Mountain Lakes, New Jersey, Yhdysvallat), joka suljettiin tyhjiörasvalla, kuten aiemmin on kuvattu (Harris et al., 2010). GFP-merkityt WT MR-1-, ΔcymA-, Δmcp_cache- ja ΔmtrBC/omcA-solut valkaistiin käyttämällä käännetyn Leica TCS SPE konfokaalimikroskoopin (Wetzlar, Saksa) maksimivalon intensiteettiasetuksia 60X-linsseillä 15 minuutin ajan. Sen varmistamiseksi, että valkaisu tapahtui, aikakuvaus kuvasi ruutualueen (250 × 250 μm valittu alue) minuutin välein, kunnes alkuperäiset solut näyttivät tummilta ja ympäröivät solut pysyivät kirkkaasti fluoresoivina. Tämän jälkeen kuvat otettiin käyttäen 588 nm:n eksitaatiota ja 530 nm:n emissiota. Time-lapse-video koko putken osasta kuvattiin käyttämällä Leica Imaging -ohjelmistoa ja ”automaattitarkennus”-ominaisuutta seuraavien 3 tunnin ajan. Soluja tarkkailtiin myös läpivalaisumoodissa sen varmistamiseksi, että valkaistut solut olivat liikkuvia ja ehjiä. Erillinen negatiivinen kontrolli, jossa oli GFP-merkitty Δmcp_cache, kuvattiin 3 tunnin ajan. Tässä negatiivisessa kontrollissa ei havaittu solujen kerääntyvän pimeään vyöhykkeeseen eikä valkaistujen solujen GFP-fluoresenssi palautunut. Koko kapillaarin (korkeus 30 μm) vaste kuvattiin aikajaksokuvauksella, ja solujen summa kaikissa 20 z-akselipinossa määritettiin (käyttäen jäljempänä esitettyä tietokoneanalyysimenetelmää) kullekin aikapisteelle.
Solujen liikkeiden mikroskooppinen talteenotto
Bakteerien seuranta- ja analyysimenetelmät olivat identtiset aiempien tutkimusten kanssa, joita hyödynnettiin kuviossa 3 ja jotka on kirjattu taulukkoon 3 (Harris et al., 2012). Sekä tietokoneella että manuaalisesti tehdyt bakteerien uintiradat standardoitiin käyttäen mittakaavaviivaa (100 μm). Kustakin kokeesta tallennettiin yleinen uintiaktiivisuus videokuvassa – joka vastaa 107 × 193 μm:n näkökenttää – ja video normalisoitiin ajallisesti, jotta saatiin uintinopeudet μm/s. Kustakin bakteerien uintiradasta mitattiin useita parametreja, kuten liikkunut kokonaismatka, radan kesto bakteerien ensimmäisestä ilmestymisestä ja katoamisesta, käännösten lukumäärä, kunkin käännöksen ja metallioksidin välinen etäisyys sekä metallioksidin ja bakteeriradan alkupisteen välinen etäisyys.
Uintianalyysi
Solut seurattiin käsin videodatasta (30 kuvaa/s), ruutu kerrallaan, ja ne mitattiin kuva-analyysiohjelmistolla ImageJ. Syötettyihin tietoihin sisältyvät uintinopeus, bakteerien lähtöasento lähimpään IEA-pintaan nähden ja jokaisen bakteerin kääntötapahtuman sijainti tunnistettiin ja kirjattiin ylös suhteessa etäisyyteen lähimmästä IEA-pinnasta (kuva 3 ja taulukko 3). Uintiaktiivisuuden määrittämiseksi sen jälkeen, kun solut olivat olleet kosketuksissa IEA:n kanssa, uivat solut jaettiin analyysia varten kahteen ryhmään: soluja, jotka uivat 2 μm:n etäisyydellä hiukkasesta, pidettiin ”kosketuksissa olevina” ja soluja, jotka eivät uineet 2 μm:n etäisyydellä hiukkasen pinnasta, pidettiin ”ei-kosketuksissa olevina”. Edellä kuvattujen käsinseurantamenetelmien lisäksi kokeelliset tiedot lajiteltiin tietokoneella ja analysoitiin aiemmin kuvatulla algoritmilla, jonka avulla saatiin lasketut uintitiedot (Crocker ja Grier, 1996; Harris et al., 2012; Harris, 2013). Kuvien 3, 4 tuottamiseksi kaikki kokeelliset tiedot neljästä erillisestä biologisesta toistosta yhdistettiin, koottiin yhteen ja keskiarvoistettiin lokeroihin etäisyyden mukaan IEA-hiukkasesta. Koska nämä tiedot yhdistettiin etäisyyksien mukaan, virhepalkit ovat päällekkäisiä; kantojen välillä ei voida tehdä vertailua käänteisfrekvenssistä, tietyllä etäisyydellä, koska tämä visualisointimenetelmä sulkee pois kokeiden tunnistetiedot.
Tekijän myötävaikutukset
HH: suunnitteli ja suoritti kokeet; HH ja IS-A: kirjoittivat ja tarkistivat paperin; ES, ME-N ja JM: auttoivat kokeiden suunnittelussa; WT: auttoivat suorittamaan data-analyysin; JM ja KN: tarkensivat käsikirjoitustaan.
Kiinnostusristiriitoja koskeva lausunto
Tekijät ilmoittavat, että tutkimus tehtiin ilman kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdolliseksi eturistiriidaksi.
Kiitokset
Erityiset kiitokset Mandy J. Wardille ja Gijs Kuenenille neuvoista tutkimuksen ja kokeiden suunnittelussa. Kiitämme Meaghan Sullivania manuaalisista seuranta-analyyseistä. Kiitämme Cecile Jourlin-Castellia, Samantha Reediä, Jun Li:tä ja David Culleya Δmcp_cache-, ΔmtrBC/omcA- ja ΔcymA-mutanttien toimittamisesta. Tämän työn rahoitus saatiin ilmavoimien tieteellisen tutkimuksen toimiston myöntämästä palkinnosta. IS-A:n tutkimusta tuki NASA:n Planetary Biology Internship -apuraha.
Supplementary Material
Tämän artikkelin Supplementary Material löytyy verkosta osoitteesta: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02568/full#supplementary-material
Alexandre, G., Greer-Phillips, S., ja Zhulin, I. B. (2004). Energiataksien ekologinen rooli mikro-organismeissa. FEMS Microbiol. Rev. 28, 113-126. doi: 10.1016/j.femsre.2003.10.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Baraquet, C., Théraulaz, L., Iobbi-Nivol, C., Méjean, V., and Jourlin-Castelli, C. (2009). Odottamattomat kemoreseptorit välittävät energian taksonointia kohti elektroniakseptoreita Shewanella oneidensis -bakteerissa. Mol. Microbiol. 73, 278-290. doi: 10.1111/j.1365-2958.2009.06770.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Beliaev, A. S., and Saffarini, D. A. (1998). Shewanella putrefaciens mtrB koodaa ulkokalvoproteiinia, jota tarvitaan Fe(III) ja Mn(IV) pelkistämiseen. J. Bacteriol. 180, 6292-6297.
PubMed Abstract | Google Scholar
Bencharit, S., ja Ward, M. J. (2005). Shewanella oneidensis MR-1:n kemotaktiset vasteet metalleille ja anaerobisille elektroniakseptoreille. J. Bacteriol. 187, 5049-5053. doi: 10.1128/JB.187.14.5049-5053.2005
PubMed Abstract | CrossRef Full Text | Google Scholar
Bond, D. R., and Lovley, D. R. (2003). Elektrodiin kiinnitetyn Geobacter sulfurreducensin sähköntuotanto. Appl. Environ. Microbiol. 69, 1548-1555. doi: 10.1128/AEM.69.3.1548-1555.2003
PubMed Abstract | CrossRef Full Text | Google Scholar
Bowman, J. P., McCammon, S. A., Nichols, D. S., Skerratt, J. H., Rea, S. M., Nichols, P. D., ym. et al. (1997). Shewanella gelidimarina sp. nov. ja Shewanella frigidimarina sp. nov., uudet antarktiset lajit, jotka kykenevät tuottamaan eikosapentaeenihappoa (20:5ω3) ja kasvamaan anaerobisesti dissimilatorisen Fe(III)-pelkistyksen avulla. Int. J. Syst. Bacteriol. 47, 1040-1047. doi: 10.1099/00207713-47-4-1040
PubMed Abstract | CrossRef Full Text | Google Scholar
Bretschger, O. (2008). Shewanella-suvun elektroninsiirtokyky ja aineenvaihduntaprosessit sekä sovellukset mikrobipolttokennojen optimointiin. University of Southern California, ProQuest Dissertations Publishing, 3325185.
Google Scholar
Bretschger, O., Obraztsova, A., Sturm, C. A., Chang, I. S., Gorby, Y. A., Reed, S. B., et al. (2007). Virran tuotanto ja metallioksidipelkistys Shewanella oneidensis MR-1:n villityypin ja mutanttien avulla. Appl. Environ. Microbiol. 73:7003. doi: 10.1128/AEM.01087-07
PubMed Abstract | CrossRef Full Text | Google Scholar
Brettar, I., Christen, R., ja Höfle, M. G. (2002). Shewanella denitrificans sp nov., voimakkaasti denitrifioiva bakteeri, joka on eristetty Itämeren keskiosassa sijaitsevan Gotlannin syvänteen hapellis-anoksisesta rajapinnasta. Int. J. Syst. Evol. Microbiol. 52, 2211-2217. doi: 10.1099/00207713-52-6-2211
PubMed Abstract | CrossRef Full Text | Google Scholar
Burdige, D. J. (1993). Mangaanin ja raudan pelkistymisen biogeokemia merisedimenteissä. Earth Sci. Rev. 35, 249-284. doi: 10.1016/0012-8252(93)90040-E
CrossRef Full Text | Google Scholar
Champine, J. E., Underhill, B., Johnston, J. M., Lilly, W. W. ja Goodwin, S. (2000). Elektroninsiirto dissimiloituvassa rautaa pelkistävässä Geobacter metallireducens -bakteerissa. Anaerobe 6, 187-196. doi: 10.1006/anae.2000.0333
CrossRef Full Text | Google Scholar
Childers, S. E., Ciufo, S., and Lovley, D. R. (2002). Geobacter metallireducens pääsee liukenemattomaan Fe(III)-oksidiin kemotaksiksen avulla. Nature 416, 767-769. doi: 10.1038/416767a
PubMed Abstract | CrossRef Full Text | Google Scholar
Coursolle, D., ja Gralnick, J. A. (2010). Shewanella oneidensis -kannan MR-1:n Mtr-hengitysreitin modulaarisuus. Mol. Microbiol. 77, 995-1008. doi: 10.1111/j.1365-2958.2010.07266.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Crocker, J. C., ja Grier, D. G. (1996). Digitaalisen videomikroskopian menetelmät kolloiditutkimuksissa. J. Colloid Interface Sci. 179, 298-310. doi: 10.1006/jcis.1996.0217
CrossRef Full Text | Google Scholar
El-Naggar, M. Y., Wanger, G., Leung, K. M., Yuzvinsky, T. D., Southam, G., Yang, J., et al. (2010). Sähkönkuljetus Shewanella oneidensis MR-1:n bakteerin nanolankoja pitkin. Proc. Natl Acad. Sci. U.S.A. 107, 18127-18131. doi: 10.1073/pnas.1004880107
PubMed Abstract | CrossRef Full Text | Google Scholar
Fredrickson, J. K., Romine, M. F., Beliaev, A. S., Auchtung, J. M., Driscoll, M. E., Gardner, T. S., et al. (2008). Towards environmental systems biology of Shewanella. Nat. Rev. Microbiol. 6, 592-603. doi: 10.1038/nrmicro1947
PubMed Abstract | CrossRef Full Text | Google Scholar
Fredrickson, J. K., Zachara, J. M., Kennedy, D. W., Dong, H., Onstott, T. C., Hinman, N. W., et al. (1998). Biogeeninen raudan mineralisaatio, joka liittyy pohjaveden bakteerin suorittamaan vesipitoisen rautaoksidin dissimilaatiopelkistykseen. Geochim. Cosmochim. Acta 62, 3239-3257. doi: 10.1016/S0016-7037(98)00243-9
CrossRef Full Text | Google Scholar
Gao, H., Obraztova, A., Stewart, N., Popa, R., Fredrickson, J. K., Tiedje, J. M., et al. (2006). Shewanella loihica sp. nov., eristetty Tyynen valtameren rautapitoisista mikrobimatoista. Int. J. Syst. Evol. Microbiol. 56, 1911-1916. doi: 10.1099/ijs.0.64354-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Gorby, Y. A., Beveridge, T. J., ja Wiley, W. R. (2005). ”Dissimilatoristen metallia pelkistävien bakteerien tuottamien solunulkoisten metallia pelkistävien rakenteiden (nanolankojen) koostumus, reaktiivisuus ja säätely”, in Annual NABIR PI Meeting (Warrenton, VA).
Google Scholar
Gorby, Y. A., Yanina, S., McLean, J. S., Rosso, K. M., Moyles, D., Dohnalkova, A., et al. (2006). Shewanella oneidensis-kannan MR-1 ja muiden mikro-organismien tuottamat sähköä johtavat bakteerien nanolangat. Proc. Natl. Acad. Sci. U.S.A. 103, 11358-11363. doi: 10.1073/pnas.0604517103
PubMed Abstract | CrossRef Full Text | Google Scholar
Gross, B. J., and El-Naggar, M. Y. (2015). Yhdistetty sähkökemiallinen ja optinen ansastusalusta yksittäisten solujen hengitysnopeuksien mittaamiseen elektrodien rajapinnoilla. Rev. Sci. Instrum. 86:064301. doi: 10.1063/1.4922853
PubMed Abstract | CrossRef Full Text | Google Scholar
Harris, H. W. (2013). Uuden bakteerien tunnistamismekanismin tunnistaminen: Bacterial Insoluble Electron Acceptor Sensingin karakterisointi. University of Southern California.
Google Scholar
Harris, H. W., El-Naggar, M. Y., Bretschger, O., Ward, M. J., Romine, M. F., Obraztsova, A. Y., et al. (2010). Elektrokinesis on mikrobien käyttäytyminen, joka edellyttää solunulkoista elektronin kuljetusta. Proc. Natl. Acad. Sci. U.S.A. 107, 326-331. doi: 10.1073/pnas.0907468107
PubMed Abstract | CrossRef Full Text | Google Scholar
Harris, H. W., Naggar, M. Y., and Nealson, K. H. (2012). Shewanella oneidensis MR-1:n kemotaksisproteiinit ja elektroninkuljetusketjun komponentit, jotka ovat välttämättömiä kokoontumiselle lähellä liukenemattomia elektroniakseptoreita. Biochem. Soc. Trans. 40, 1167-1177. doi: 10.1042/BST20120232
PubMed Abstract | CrossRef Full Text | Google Scholar
Hau, H. H. ja Gralnick, J. A. (2007). Shewanella-suvun ekologia ja bioteknologia. Annu. Rev. Microbiol. 61, 237-258. doi: 10.1146/annurev.micro.61.080706.093257
PubMed Abstract | CrossRef Full Text | Google Scholar
Hsu, L., Masuda, S. A., Nealson, K. H., ja Pirbazari, M. (2012). Mikrobipolttokennon Shewanella-biokatodien arviointi kromaattisaasteen käsittelyssä. R. Soc. Chem. Adv. 2, 5844-5855. doi: 10.1039/c2ra20478a
CrossRef Full Text | Google Scholar
Kan, J., Hsu, L., Cheung, A. C. M., Pirbazari, M. ja Nealson, K. H. (2011). Bakteeriyhteisöjen virran tuotanto mikrobipolttokennoissa, jotka on rikastettu jätevesilietteestä eri elektroninluovuttajilla. Environ. Sci. Technol. 45, 1139-1146. doi: 10.1021/es102645v
PubMed Abstract | CrossRef Full Text | Google Scholar
Kato, S., Nakamura, R., Kai, F., Watanabe, K. ja Hashimoto, K. (2010). Maaperän bakteerien hengitysvuorovaikutukset (puoli)johtavien rautaoksidimineraalien kanssa. Environ. Microbiol. 12, 3114-3123. doi: 10.1111/j.1462-2920.2010.02284.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Kotloski, N. J., ja Gralnick, J. A. (2013). Flaviinielektronisukkulat hallitsevat Shewanella oneidensiksen solunulkoista elektroninsiirtoa. MBio 4:e00553. doi: 10.1128/mBio.00553-12
PubMed Abstract | CrossRef Full Text | Google Scholar
Kracke, F., Vassilev, I., and Krömer, J. O. (2015). Mikrobien elektroninsiirto ja energiansäästö – perusta bioelektrokemiallisten järjestelmien optimoinnille. Front. Microbiol. 6:575. doi: 10.3389/fmicb.2015.00575
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, J., Go, A. C., Ward, M. J., ja Ottemann, K. M. (2010). Kemikaali-pistokkeessa oleva bakteerien kemotaksismääritys on altis väärille positiivisille vasteille. BMC Res. Notes 3:77. doi: 10.1186/1756-0500-3-77
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, J., Romine, M. F., ja Ward, M. J. (2007). Erittäin konservoituneen kemotaksisgeeniklusterin tunnistaminen ja analysointi Shewanella-lajeissa. FEMS Microbiol. Lett. 273, 180-186. doi: 10.1111/j.1574-6968.2007.00810.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, R., Tiedje, J. M., Chiu, C., ja Worden, R. M. (2012). Liukoiset elektronisukkulat voivat välittää energiataksoja kohti liukenemattomia elektroniakseptoreita. Environ. Sci. Technol. 46, 2813-2820. doi: 10.1021/es204302w
PubMed Abstract | CrossRef Full Text | Google Scholar
Lovley, D. R., Coates, J. D., Blunt-Harris, E. L., Phillips, E. J. P., ja Woodward, J. C. (1996). Humusaineet mikrobihengityksen elektroniakseptoreina. Nature 382, 445-448. doi: 10.1038/382445a0
CrossRef Full Text | Google Scholar
Lovley, D. R., Giovannoni, S. J., White, D. C., Champine, J. E., Phillips, E. J., Gorby, Y. A., et al. (1993). Geobacter metallireducens gen. nov. sp. nov., mikro-organismi, joka kykenee yhdistämään orgaanisten yhdisteiden täydellisen hapettumisen raudan ja muiden metallien pelkistämiseen. Arch. Microbiol. 159, 336-344. doi: 10.1007/BF00290916
PubMed Abstract | CrossRef Full Text | Google Scholar
Lovley, D. R., Holmes, D. E., and Nevin, K. P. (2004). Dissimilatorinen Fe(III) ja Mn(IV) pelkistyminen. Adv. Microb. Physiol. 49, 219-286. doi: 10.1111/j.1462-2920.2004.00593.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Marsili, E., Baron, D. B., Shikhare, I. D., Coursolle, D., Gralnick, J. A., Bond, D. R., et al. (2008). Shewanella erittää flaviineja, jotka välittävät solunulkoista elektroninsiirtoa. Proc. Natl. Acad. Sci. U.S.A. 105, 3968-3973. doi: 10.1073/pnas.0710525105
PubMed Abstract | CrossRef Full Text | Google Scholar
McLean, J. S., Majors, P. D., Reardon, C. L., Bilskis, C. L., Reed, S. B., Romine, M. F., ym. et al. (2008). Shewanella oneidensis MR-1 -biofilmien rakenteen ja aineenvaihdunnan tutkiminen. J. Microbiol. Methods 74, 47-56. doi: 10.1016/j.mimet.2008.02.015
PubMed Abstract | CrossRef Full Text | Google Scholar
Meyer, T. E., Tsapin, A. I., Vandenberghe, I., de Smet, L., Frishman, D., Nealson, K. H., ym. et al. (2004). 42 mahdollisen sytokromi C -geenin tunnistaminen Shewanella oneidensis -genomissa ja kuuden liukoisen sytokromin karakterisointi. Omics 8, 57-77. doi: 10.1089/153623104773547499
PubMed Abstract | CrossRef Full Text | Google Scholar
Mitchell, A. C., Peterson, L., Reardon, C. L., Reed, S. B., Culley, D. E., Romine, M. R., et al. (2012). Ulkokalvon c-tyypin sytokromien MtrC ja OmcA rooli Shewanella oneidensis MR-1 -solujen tuotannossa, kerääntymisessä ja irtoamisessa hengityksen aikana hematiitilla. Geobiology 10, 355-370. doi: 10.1111/j.1472-4669.2012.00321.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Murray, A. E., Lies, D., Li, G., Nealson, K., Zhou, J., ja Tiedje, J. M. (2001). DNA/DNA-hybridisaatio mikrosarjoihin paljastaa geenispesifisiä eroja läheisesti sukua olevien mikrobien genomien välillä. Proc. Natl. Acad. Sci. U.S.A. 98, 9853-9858. doi: 10.1073/pnas.171178898
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Myers, J. M. (1997). Cloning and sequence of cymA, a gene encoding a tetraheme cytochrome c required for reduction of iron(III), fumarate, and nitrate by Shewanella putrefaciens MR-1 . J. Bacteriol. 179, 1143-1152. doi: 10.1128/jb.179.4.1143-1152.1997
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Myers, J. M. (2002). MtrB tarvitaan sytokromien OmcA ja OmcB asianmukaiseen sisällyttämiseen Shewanella putrefaciens MR-1:n ulkokalvoon. Appl. Environ. Microbiol. 68, 5585-5594. doi: 10.1128/AEM.68.11.5585-5594.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Nealson, K. H. (1988a). Bakteerien mangaanin pelkistyminen ja kasvu mangaanioksidilla ainoana elektroniakseptorina. Science 240, 1319-1321. doi: 10.1126/science.240.4857.1319
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Nealson, K. H. (1988b). Mangaanioksidien mikrobiologinen pelkistäminen: Vuorovaikutukset raudan ja rikin kanssa. Geochim. Cosmochim. Acta 52, 2727-2732. doi: 10.1016/0016-7037(88)90041-5
CrossRef Full Text | Google Scholar
Myers, J. M., and Myers, C. R. (2001). Shewanella putrefaciens MR-1:n ulkokalvon sytokromien OmcA ja OmcB rooli mangaanidioksidin pelkistämisessä. Appl. Environ. Microbiol. 67, 260-269. doi: 10.1128/AEM.67.1.260-269.2001
PubMed Abstract | CrossRef Full Text | Google Scholar
Nealson, K. H., Moser, D. P., and Saffarini, D. A. (1995). Anaerobisen elektronin akseptorin kemotaksis Shewanella putrefaciensissa. Appl. Environ. Microbiol. 61, 1551-1554.
PubMed Abstract | Google Scholar
Okamoto, A., Kalathil, S., Deng, X., Hashimoto, K., Nakamura, R., ja Nealson, K. H. (2014). Kalvon sytokromeihin sitoutuneet solun erittämät flaviinit sanelevat elektroninsiirtoreaktioita pinnoille, joilla on erilainen varaus ja pH. Sci. Rep. 4:5628. doi: 10.1038/srep05628
PubMed Abstract | CrossRef Full Text | Google Scholar
Pirbadian, S., Barchinger, S. E., Leung, K. M., Byun, H. S., H. S., Jangir, Y., Bouhenni, R. A., et al. (2015). Shewanella oneidensis MR-1:n bakteerin nanolangat ovat solunulkoisten elektroninkuljetuskomponenttien ulkokalvon ja periplasman jatkeita. Biophys. J. 108:368a. doi: 10.1016/j.bpj.2014.11.2016
CrossRef Full Text | Google Scholar
Reguera, G., McCarthy, K. D., Mehta, T., Nicoll, J. S., Tuominen, M. T., Lovely, D. R., et al. (2005). Solunulkoinen elektroninsiirto mikrobiologisten nanolankojen avulla. Nature 435, 1098-1101. doi: 10.1038/nature03661
PubMed Abstract | CrossRef Full Text | Google Scholar
Salas, E. C., Berelson, W. M., Hammond, D. E., Kampf, A. R. ja Nealson, K. H. (2010). Bakteerikannan vaikutus dissimilatorisen raudan pelkistymisen tuotteisiin. Geochim. Cosmochim. Acta 74, 574-583. doi: 10.1016/j.gca.2009.10.039
PubMed Abstract | CrossRef Full Text | Google Scholar
Saltikov, C. W., Cifuentes, A., Venkateswaran, K. ja Newman, D. K. (2003). Ars-detoksifikaatiojärjestelmä on edullinen mutta ei välttämätön geneettisesti käsiteltävän Shewanella-lajin kannan ANA-3 As(V)-hengitykselle. Appl. Environ. Microbiol. 69, 2800-2809. doi: 10.1128/AEM.69.5.2800-2809.2003
CrossRef Full Text | Google Scholar
Schwalb, C., Chapman, S. K., and Reid, G. A. (2003). Tetrahemisytokromi CymA:ta tarvitaan anaerobiseen hengitykseen dimetyylisulfoksidin ja nitriitin kanssa Shewanella oneidensis -bakteerissa. Biochemistry 42, 9491-9497. doi: 10.1021/bi034456f
PubMed Abstract | CrossRef Full Text | Google Scholar
Schwertmann, U., ja Cornell, R. M. (2008). Rautaoksidit laboratoriossa. Weinheim: John Wiley & Sons.
Google Scholar
Shi, L., Richardson, D. J., Wang, Z., Kerisit, S. N., Rosso, K. M., Zachara, J. M., et al. (2009). Shewanellan ja Geobacterin ulkokalvon sytokromien rooli solunulkoisessa elektroninsiirrossa. Environ. Microbiol. Rep. 1, 220-227. doi: 10.1111/j.1758-2229.2009.00035.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Venkateswaran, K., Dollhopf, M. E., Aller, R., Stackebrandt, E. ja Nealson, K. H. (1998). Shewanella amazonensis sp. nov., uusi metallia pelkistävä fakultatiivinen anaerobi Amazonin mannerjalustamudasta. Int. J. Syst. Bacteriol. 48, 965-972.
PubMed Abstract | Google Scholar
Venkateswaran, K., Moser, D. P., Dollhopf, M. E., Lies, D. P., Saffarini, D. A., MacGregor, B. J., et al. (1999). Shewanella-suvun polyfaattinen taksonomia ja Shewanella oneidensis sp. nov:n kuvaus. Int. J. Syst. Bacteriol. 49, 705-724. doi: 10.1099/00207713-49-2-705
PubMed Abstract | CrossRef Full Text | Google Scholar
Ziemke, F., Höfle, M. G., Lalucat, J. ja Rossellö-Mora, R. (1998). Shewanella putrefaciens Owenin genomiryhmän II uudelleenluokittelu Shewanella baltica sp. nov. Int. J. Syst. Bacteriol. 48, 179-186. doi: 10.1099/00207713-48-1-179
PubMed Abstract | CrossRef Full Text | Google Scholar