Introduction
Luonnollisten korjaustoimenpiteiden käyttö on ikivanha perinne, joka on edelleen käytössä, ja >60 % maailman väestöstä käsittelee erilaisia terveysongelmia, mukaan lukien syöpä, käyttämällä perinteistä lääketiedettä ensimmäisenä valintana (Sultana et al., 2014). Syövän hoito on kuitenkin monimutkaista, ja potilaiden nykyiset mahdollisuudet riippuvat syövän tyypistä ja vaiheesta sekä potilaan iästä, sukupuolesta ja yleisestä terveydentilasta. Vaikka solunsalpaajahoito onnistuu yleensä syövän alkuvaiheessa, sen teho riippuu lääkkeen annosteluohjelmasta ja potilaan fysiologisesta tilasta. Suurin huolenaihe kemoterapian suhteen on kuitenkin sen myrkyllisyys, sillä käytetyt lääkkeet vaikuttavat yleensä sekä syöpäsoluihin että normaaleihin soluihin, ja korkea proliferaatioindeksi aiheuttaa potilaille liitännäisvaurioita. Lisäksi syöpäsolut voivat kehittää hoitoresistenssiä ja saada etäpesäkkeitä, mikä heikentää hoitovastetta ja vähentää taudista vapaan eloonjäämisen mahdollisuutta. Siksi yksi lääkekehityksen keskeisistä haasteista on kemoterapeuttisten aineiden toksisuuden vähentäminen ja tehokkaampien ja tehokkaampien lääkkeiden kehittäminen hoitojen, toipumisaikojen ja potilaiden yleisen elämänlaadun parantamiseksi.
Luonnontuotteet ovat tarjonneet runsaan kemiallisten rakenteiden lähteen syövänvastaisten hoitojen kehittämistä varten. Syöpälääketieteen alalla 49 prosenttia kemoterapiassa käytetyistä lääkkeistä on peräisin luonnollisista lähteistä, kuten kasveista, mikro-organismeista ja meren eliöistä, tai niiden innoittamina (Newman ja Cragg, 2016). Esimerkkeinä voidaan mainita vinca-alkaloidit ja taksaanit (tubuliinia sitovat aineet) sekä podofyllotoksiinit, antrasykliinit ja etoposidit (topoisomeraasin estäjät) (Da Rocha ym., 2001; Nobili ym., 2009). Nämä esimerkit havainnollistavat luonnontuotteiden potentiaalia lääkkeiden löytämisessä.
AA:t ovat Annonaceae-suvun tuottamia sekundaarisia aineenvaihduntatuotteita (kuva 1). Perinteisessä lääketieteessä Annona-suvun hedelmiä käytetään kuumeen, kivun, reumatismin, ripulin ja niveltulehduksen hoitoon ja sen lehtiä diabeteksen, päänsäryn ja unettomuuden hoitoon (Moghadamtousi et al., 2015a). Annona reticulatea on käytetty Afrikassa dysenteriakipulääkkeenä ja heminttilääkkeenä, ja A. squamosaa käytetään Intiassa erilaisten sairauksien, kuten pahanlaatuisten kasvainten, hoitoon (Savithramma et al., 2011). Lisäksi A. muricata on suosittu lääke Amerikassa, Afrikassa ja Intiassa syövän hoitoon (Moghadamtousi et al., 2015a), kun taas Meksikossa A. macroprophyllata, A. muricata ja A. purpurea käytetään ihokasvainten ja mahasyövän hoitoon (Alonso-Castro et al., 2011; Brindis et al., 2013). Meksikon lääkekasveja koskevan etnofarmakologisen katsauksen mukaan Annona-sukua voitaisiin tutkia myös paksusuolen syövän hoidossa (Jacobo-Herrera et al., 2016). Nämä todisteet osoittavat, että AA:t ovat molekyylejä, joilla on merkittävää bioaktiivista potentiaalia.

KUVA 1 Asetogeniinien ytimet: lineaali, epoksi, mono-tetrahydrofuraani (THF), bis-THF ja tris-THF.
Tähän mennessä on kuvattu yli 500 AA:ta; näille yhdisteille on ominaista pitkä alifaattinen ketju, jossa on α, β-tyydyttymätön γ-laktonirengas ja 0-3 tetrahydrofuraani- (THF-) rengasta (Yuan ym, 2015), ja ne ovat osoittaneet sytotoksista aktiivisuutta erilaisia ihmisen syöpäsolulinjoja (eli keuhko-, rinta-, paksusuoli-, munuais-, haima-, eturauhas-, maksa- ja virtsarakkosolulinjoja) vastaan (katsaus artikkelissa Zeng et al., 1996; Chávez ja Mata, 1998; Wang et al., 2002; Yuan et al., 2003; Mangal et al., 2016; Miao et al., 2016a; Miao et al., 2016b) lisäksi in vivo kasvainvastainen aktiivisuus (Chen et al., 2012a; Chen et al., 2012b). Tässä tarkastelimme viimeisimpiä AA:ta koskevia raportteja lääkekeksintö-onkologian alalla.
Materiaalit ja menetelmät
Toteutimme haun PubMed-tietokannasta käyttäen seuraavia hakusanoja: ”acetogenins and cancer”, ”acetogenins antitumor activity”, ”acetogenins cytotoxicity” ja ”acetogenins mechanism of action”, muiden yhdistelmien ohella. Tarkastelimme viimeisten 15 vuoden aikana julkaistuja artikkeleita, jotka perustuivat kahteen kriteeriin: lääkinnälliseen käyttöön tarkoitetut kasvit ja biotutkimukseen perustuva fraktiointimenetelmä. Tässä raportoidut in vitro -tiedot perustuvat yksinomaan puhtaiden yhdisteiden aktiivisuuteen, kun taas in vivo -tutkimuksissa otettiin huomioon myös kasviuutteet, koska eläinmalleissa tehtyjä kokeita ei ollut saatavilla.
Tulokset ja keskustelu
In vitro -tutkimukset
Luonnontuotealalla bioaktiivisten molekyylien etsimiseen liittyy yleensä uutteiden biotutkimusohjattu fraktiointi, jonka avulla pyritään etsimään aktiivisuudesta vastuussa oleva pääasiallinen tai olevat pääasialliset molekyylit. Monet sekundaariset aineenvaihduntatuotteet voivat kuitenkin kohdistua useisiin syövän tunnusmerkkeihin. Yleisesti ottaen lääketeollisuus on kiinnostunut lääkkeistä, joilla on useampi kuin yksi molekyylivuorovaikutus ja jotka kattavat erilaisia molekyylikohteita.
Acetogeniinit ovat molekyylejä, joilla on suuri potentiaali tulevaisuuden syöpähoidossa. Niiden merkittävin biologinen aktiivisuus on mitokondriokompleksi I:n esto, joka johtuu niiden bis-THF-rakenteesta. Aiemmin raportoitiinkin, että mono-THF-AA:t, joissa on alkyyliketju, joka yhdistää laktoniosan THF-ryhmään, ovat hengitysketjun kompleksi I:n (eli NADH: ubikinoni-oksidoreduktaasin) ei-kilpailukykyisiä estäjiä, mikä johtaa fosforylatiivisen hapettumisen estymiseen ja sitä seuraavaan ATP:n tuotannon vähenemiseen (Tormo ym., 1999; Chen ym., 2012). Tällainen esto koskee suurta joukkoa reittejä, jotka voivat aiheuttaa solukuolemaa, mukaan lukien apoptoosi ja autofagia, tai toimia muissa aineenvaihduntaverkoissa laktaattidehydrogenaasi A -entsyymin estäjinä, antioksidanttina tai pysäyttämällä solusyklin.
Lisäksi A. muricata (Moghadamtousi ym, 2015a) ja A. squamosa L. (Chen et al., 2012a; Chen et al., 2012b; Chen et al., 2012c; Chen et al., 2016) on raportoitu olevan sytotoksisia useisiin ihmisen tai muiden nisäkkäiden syöpäsolulinjoihin. Erityisesti AA:t annoskvatiinit A (1) ja B (2) osoittavat selektiivisyyttä MCF-7- ja A-549-solulinjoille (Chen et al., 2012c). Taulukossa 1 esitetään viimeisimmät löydetyt AA:t, joilla on sytotoksinen aktiivisuus, sekä muut vähemmän tuoreet yhdisteet, joilla on antineoplastista aktiivisuutta in vivo (taulukko 2).
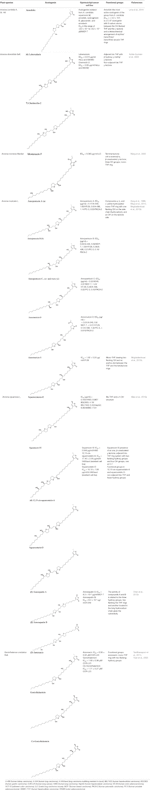
Taulukko 1 Asetogeniinien sytotoksinen aktiivisuus in vitro eri syöpäsolulinjoissa.

Taulukko 2 Asetogeniinien ja valittujen runsaasti asetogeniinejä sisältävien uutteiden kasvainvastainen aktiivisuus in vivo.
Apoptoosi on luonnollinen solukuolemastrategia, joka tappaa tarpeettomat tai vaurioituneet solut. Tärkeimmät tähän prosessiin osallistuvat geenit ovat p53 ja bcl2-perhe; edellinen on kasvainsuppressori, kun taas jälkimmäiset voivat olla joko proapoptoottisia (muun muassa BAD, BAX ja BAK) tai antiapoptoottisia (bcl2 ja bcl-x) (Okada ja Mak, 2004). P53:n ensisijainen tehtävä on estää DNA-vaurioituneiden solujen monistuminen. Näin ollen p53 on inaktiivinen, ja vaurioituneet solut jatkavat kasvuaan ja monistavat DNA-mutaatioita, mikä johtaa sairauksiin, kuten syöpään (Igney ja Krammer, 2002; Okada ja Mak, 2004). Tähän mennessä tehty tutkimus voidaankin jakaa kahteen pääaiheeseen: apoptoosin indusointiin tai apoptoosiresistenssimekanismeihin.
Nykyaikaiset apoptoosia indusoivat kemoterapiat aiheuttavat potilaille vakavia sivuvaikutuksia. Siksi vähemmän toksisten lääkkeiden etsiminen on ensisijaisen tärkeää, ja luontaistuotteiden odotetaan auttavan apoptoosia moduloivien lääkkeiden kehittämisessä. Tässä yhteydessä AA:t säätelevät kaspaasi 3:n ja 8:n (apoptoosivaikuttajat) aktiivisuutta ja vähentävät samalla survivinin ja Bcl-2:n ilmentymistä, mikä tehostaa apoptoosia. AA-annonasiini (3) edistää apoptoosia syöpäsoluissa aktivoimalla kaspaasi 3:n ja Baxin reittejä (Yuan ym., 2003), kun taas squamocin indusoi apoptoosia proapoptoottisten Bax- ja Bad-geenien ilmentymisen kautta, mikä johtaa PARP:n pilkkoutumiseen ja kaspaasi 3:n lisääntyneeseen aktiivisuuteen virtsarakon T24-syövän soluissa (Yuan ym., 2006). Tämä on ristiriidassa aiempien tulosten kanssa, joissa squamocin ei aiheuttanut apoptoosia rintasyöpäsoluissa, mutta esti proliferaatiota estämällä solusyklin G1-vaiheessa (Raynaud ym., 1999).
A. reticulatan metanoliuute estää kaspaasi 6:n ja 9:n ilmentymistä paksusuoli- ja maksasyöpäsoluissa (Mondal et al., 2007), kun taas A. squamosan orgaaniset ja vesipitoiset uutteet alentavat Bcl-2:n ilmentymistä MCF-7-rintasyöpäsoluissa ja K-562-leukemiasoluissa, mikä viittaa niiden vaikutukseen apoptoosin indusoijina (Pardhasaradhi et al., 2005). Lisäksi A. muricatan lehtiuute indusoi kaspaasi 3:n ja 9:n ilmentymistä ja estää solujen proliferaatiota vähentämällä fosfo-ERK:ta ja fosfo-ATK:ta MIA PaCa-2 -soluissa (Yiallouris ym., 2018).
AA:t johtavat myös syklipysähdykseen, millä on vaikutuksia kasvainsolujen proliferaatioon. AA:t säätelevät solusykliä G1/S-siirtymässä estämällä sykliini D1:n ilmentymistä ihmisen hepatosellulaarisissa karsinoomasoluissa (Qian ym., 2015). Tässä yhteydessä A. muricata -uute pysäyttää solusyklin G1-vaiheessa ja vähentää S-vaiheessa olevien solujen määrää pitoisuusriippuvaisesti vähentämällä solusyklin D1:n, joka on tärkeä solusyklin säätelyproteiini, ilmentymistä (Torres et al., 2012). Samanlainen tulos havaittiin squamocinilla, joka pysäyttää solut G1-vaiheessa T24-virtsarakon syöpäsoluissa (Yuan ym., 2006). Huolimatta solusyklin merkityksestä vain harvat tutkimukset ovat käsitelleet sitä, miten AA:t vaikuttavat tähän mekanismiin.
Aerobinen glykolyysi, mekanismi, jota kasvainsolut käyttävät saadakseen energiaa hapen puuttuessa (Figueroa-González ym., 2016; García-Castillo ym., 2017), on myös AA:iden kohteena. Erilaisia proteiineja ja glykolyyttisiä entsyymejä säätelee ylöspäin HIF-1, joka on tärkeä kasvaimen aerobiseen glykolyysiin osallistuva transkriptiotekijä, syöpäsoluissa. Mielenkiintoista oli, että A. muricata -uute alensi HIF-1α:n ja NF-κB:n ilmentymistä sekä glukoosin kuljettajan GLUT1:n ja HKII- ja LDHA-entsyymien tasoja haimasyöpäsoluissa (Torres ym., 2012). Lisäksi A. muricatan lehtiuute osoitti antiproliferatiivisia vaikutuksia syöpäsolulinjoissa ja edisti solukuolemaa estämällä NKA- ja SERCA-pumppuja (Yiallouris et al., 2018).
Toisaalta MDR1-geeni koodaa solukalvon glykoproteiini P:tä, joka on keskeinen kuljettajaproteiini, joka purkaa syöpälääkkeet ulos solujen sisältä ja rajoittaa siten niiden solunsisäistä kertymistä ja vähentää niiden toksisuutta (Figueroa-González ym., 2012). Sekundaariset metaboliitit, kuten flavonolit, ginsenosidit, polyfenolit, alkaloidit ja hartsiglykosidit, ovat osoittaneet mielenkiintoisia tuloksia P-gp:n moduloinnissa syöpäsolulinjoissa (Phang et al., 1993; Beck et al., 1988; Silva et al., 2001; Jodoin et al., 2002; Zhou et al., 2004; Figueroa-González et al., 2012). Lisäksi AA:t voivat alentaa MDR1- ja MRP1-geenien ilmentymistä lääkkeille resistentissä ihmisen hepatosellulaarisessa karsinoomassa sekä topoisomeraasi IIα:n ja glutationi S-transferaasiΠ:n ilmentymistä (Qian ym., 2015).
Viime aikoina autofagia on herättänyt kiinnostusta solukuolemamekanismina. Autofagia on katabolinen prosessi, jonka eukaryoottisolut aktivoivat stressitilanteessa, kuten solujen nälkiintyessä tai patogeenien läsnä ollessa (He ja Klionsky, 2009). Tämän prosessin aikana solu kierrättää proteiineja tai ei-toimivia organelleja monivaiheisessa prosessissa, johon liittyy lysosomaalinen hajoaminen, jotta solun homeostaasi voidaan lopulta palauttaa (Kenific ja Debnath, 2015). Sinänsä autofagian virtauksen deregulaatio voi johtaa syöpään, sillä makromolekyylien, ei-toimivien organellien ja proteiinien kierrättäminen tarjoaa syöpäsoluille suuret metaboliset vaatimukset solujen lisääntymiselle (Kenific ja Debnath, 2015; García-Castillo ym., 2017). Tässä skenaariossa autofagiasta on tullut houkutteleva terapeuttinen kohde, ja AA:t saattavat pystyä estämään tätä prosessia. Liu et al. (2012) raportoivat, että yhdiste AA005 (asetogeniinianalogi) esti ATP:n tuotantoa, aktivoi AMPK:ta ja esti mTOR-kompleksi 1 -reitin indusoidakseen lopulta autofagian paksusuolen syöpäsoluissa ja pysäyttääkseen solusyklin G1-vaiheeseen. Tämä yhdiste on AA-yhdisteiden jäljitelmä, jossa etyleeniglykolieetteriyksikkö korvaa kaksi THF-rengasta. Tällainen kemiallinen modaliteetti antaa ylivoimaisen biologisen aktiivisuuden, mikä on esimerkki AA:iden plastisuudesta, jota voidaan pitää inspiraationa uusien ja tehokkaampien molekyylien luomiseksi, joilla on erilaiset farmakologiset kohteet.
Acetogeniinien syöpälääkkeiden torjunta-aktiivisuus eläinmalleissa
Tiedot AA:iden ja Annona-uutteiden syövänvastaisesta aktiivisuudesta ovat erityisen lupaavia, koska näillä yhdisteillä on havaittu syöpälääkkeiden torjunta-aktiviteettia eläimissä. In vivo -tutkimukset antavat keskeistä tietoa lääkkeen suorituskyvystä koko organismissa, ja niiden avulla voidaan arvioida paitsi niiden kasvainvastaista vaikutusta myös niiden toksisuutta eri elimissä. 2012b). Ehdotettiin, että yhden komponentin cis-konfiguraatio voisi olla vastuussa kasviperäisen valmisteen sytotoksisesta aktiivisuudesta. Lisäksi A. squamosasta eristetty bulatasiini (5) annoksella 15 μg/kg vähensi tehokkaasti kasvaimen kasvua hiirillä, joilla oli S180- ja HepS-ksenotransplantaatteja, vastaavasti 65,8 ja 63,4 %. Nämä tulokset ovat parempia kuin suuremmilla taksolipitoisuuksilla (40 μg/kg) saadut tulokset. Bullatasiinilla (5) on vierekkäinen bis-THF-osa ja kolme hydroksiryhmää, jotka todennäköisesti muodostavat bioaktiivisen rakenteen (Chen ym., 2012a).
A. squamosan siemenöljy esti 53,54 % kasvaimen kasvusta hiirillä, joilla oli H22-soluja (Chen ym., 2016), ja vähensi IL-6:n, Jak:n ja erilaisten fosforyloitujen signaalinmuuntimien ja transkription aktivaattoreiden ilmentymää p-Stat3-reittiä. On myös raportoitu, että AA:ssa esiintyvät α, β-tyydyttymättömät γ-laktoniryhmät ovat Michaelin reaktioakseptoreita (Ji et al., 2012), jotka estävät Stat3-aktivoitumista, joka on terapeuttinen kohde, joka on osallisena solujen proliferaatiossa, apoptoosissa, tulehduksessa ja angiogeneesissä (Cafferkey ja Chau, 2016). A. muricata -uute hidasti kasvaimen kasvua haiman ksenotransplanteissa (Yiallouris et al., 2018), vähentää etäpesäkkeitä vähentämällä metalloproteinaasi-9:n tasoja ja edistää syöpäsolujen kuolemaa nekroosin kautta (Torres ym., 2012). Eturauhasen ksenograftissa flavonoideilla rikastettu A. muricata -uute paransi biologista hyötyosuutta ja osoitti vähäisempää toksisuutta kuin AA-yhdisteillä rikastettu uute (Yang ym., 2015).
Laherraduriini (6) ja cherimoliini-2 (7) eristettiin lääkekasvista A. diversifolia, ja niitä testattiin in vivo kohdunkaulan- ja paksusuolen syöpäsoluja vastaan (Schlie-Guzmán ym., 2009). Molemmat AA-yhdisteet pienensivät HeLa-kasvainten kokoa samanlaisilla arvoilla kuin doksorubisiini; molemmilla yhdisteillä oli myös antiproliferatiivista aktiivisuutta in vitro samaa syöpäsolulinjaa vastaan. Nämä tulokset ovat sopusoinnussa muiden raporttien kanssa, joissa aktiivisimmaksi molekyyliksi raportoitiin laherraduriini (6), jolla on vierekkäisiä bis-THF-osia ja β-hydroksi γ-metyyli γ-laktonirakenne, kun taas kemoliini-2:lla (7), joka sisältää vierekkäistä bis-THF-osaa vailla olevan bis-THF-osan ja γ-laktoniyksikön, on vähäisempi kasvainvastainen aktiivisuus. Taulukossa 2 esitetään eri AA-yhdisteiden kasvainvaikutus eläinmalleissa.
Myrkyllisyystutkimukset
Eläinkokeisiin tulisi sisältyä akuuttia suun kautta tapahtuvaa myrkyllisyyttä koskevia protokollia kemikaalien testaamiseksi ja myrkyllisyyden merkkien havainnoimiseksi; näillä minimoidaan tarvittavien eläinten määrä, varmistetaan oikean annoksen antaminen kokeissa ja vältetään kärsimys (OECD, 2008). Toistaiseksi AA-yhdisteiden tai Annona-uutteiden myrkyllisyydestä on kuitenkin vain vähän tietoa. Esimerkiksi A. muricatan etanoliuutteen LD50-arvo hiirillä oli Sousa et al. (2010) mukaan 1,67 g/kg. Arthur et al. (2011) ilmoittivat vesiuutteen LD50-arvon (<5 g/kg) ja kirjasivat, että suurempi annos saattaa vahingoittaa munuaisia. Lisäksi havaittiin, että A. squamosan siemenuute voi aiheuttaa maksavaurioita (Miao et al., 2016b), ja flavonoideilla rikastettu A. muricata -uute osoitti vähentynyttä myrkyllisyyttä eturauhasen ksenograftissa (Yang et al., 2015). A. diversifoliasta eristetyllä cherimolin-2:lla oli myrkyllisiä vaikutuksia ja kuolema SW480-ksenotransplanteissa annoksilla, jotka olivat yli 5 mg/kg/vrk (Schlie-Guzmán ym., 2009).
Kuten aiemmin selitettiin, AA:t voivat estää hengitysketjun kompleksi 1:n elektroniketjun kuljetuksen mitokondrioissa. AA:t liittyvät ja estävät NADH-entsyymin, joka yleensä yli-ilmentyy syöpäsoluissa, estämällä ATP:n tuotannon, mikä johtaa lopulta solukuolemaan. Oletettavasti tällainen mekanismi viittaa siihen, että AA:t ovat ”vaarattomia” normaaleille soluille; silti olisi tehtävä lisää tutkimuksia näiden molekyylien selektiivisyyden varmistamiseksi.
Johtopäätös
Sytostaattihoito ei ole spesifistä syöpäsoluille, vaan se aiheuttaa useita ei-toivottuja sivuvaikutuksia, kuten vaurioita normaaleille kudoksille ja elimille. Tavanomaisen kemoterapian tärkein näkökohta on kuitenkin se, että merkittävässä osassa tapauksista syöpäsolut kehittävät resistenssimekanismeja, jotka mahdollistavat kasvaimen etenemisen ja etäpesäkkeiden muodostumisen.
Uusien lääkkeiden etsiminen luonnosta ei ole uutta. Useita luonnon aineiden johdannaisia käytetään tällä hetkellä eri sairauksien hoitoon. Esimerkiksi perimätiedon avulla löydettyä taksolia (paklitakselia) pidetään markkinoiden kannattavimpana syöpälääkkeenä. Mielenkiintoista on, että paklitakseli on antimitoottinen lääke, joka estää solujen lisääntymistä soluviljelmissä, mutta kasvaimissa sen on todettu aiheuttavan moninapaista jakautumista, mikä kuvaa sitä, miten molekyylillä voi olla erilaisia vaikutusmekanismeja riippuen in vitro- tai potilasolosuhteista. Tällä alkaloidilla on 30 vuoden historia, ja se havainnollistaa luonnontuotteilla työskentelyn suurimpia esteitä, niiden rajallista biologista saatavuutta ja tuotantokustannuksia, mutta myös uskomatonta tehoa syöpäsoluja tappaessa.
Acetogeniinit ovat monipuolisia syöpämolekyylejä, jotka aiheuttavat kasvainsolujen kuoleman eri mekanismeilla. Ne voivat moduloida kemoterapeuttisten lääkkeiden poissulkemista syöpäsoluista ja ovat voimakkaita apoptoosin indusoijia. Niiden bioaktiivinen joustavuus näkyy niiden kyvyssä säädellä solusykliä pysäyttämällä solut G1-vaiheeseen, edistää apoptoosia estämällä eri proteiineja ja jopa indusoida autofagiaa. Lisäksi niiden metaboliset vuorovaikutukset, jotka liittyvät erityisesti transkriptiotekijöihin HIF1 ja STAT3, ja niiden vaikutukset energiankulutukseen, angiogeneesiin, tulehdukseen ja metastaasiin, ovat merkittäviä. AA-yhdisteiden kasvainvastainen aktiivisuus in vivo on lupaava (esimerkkeinä bullatasiini, laherraduriini ja serimoliini-2). Prekliiniset tiedot eivät kuitenkaan riitä AA-yhdisteiden farmakodynamiikan ja kinetiikan ymmärtämiseen, ja akuuttia toksisuutta ja liukoisuutta koskevia testejä tarvitaan lisää, jotta voidaan varmistaa turvallisuus ja mahdollisuus kliinisiin tutkimuksiin ihmisillä. Lisäksi erilaisten ligandien (esim. vasta-aineiden, vitamiinien ja peptidien) sisällyttäminen tai kasvainspesifisten johdannaisten valmistaminen voisi parantaa AA:n aktiivisuutta ja tuottaa sopivampia lääkkeitä.
Kasviuutteet ovat kemiallisesti monimutkaisia, ja niiden parantavat ominaisuudet riippuvat usein yhdisteiden välisistä vuorovaikutuksista ja niiden osuuksista uutteessa. AA-yhdisteiden vaikutuksista syöpään on vielä paljon löydettävää. Tämä ala tarjoaa mahdollisuuden löytää uusia molekyylejä tämän monimutkaisen sairauden hoitoon. Uutteiden standardointi on näin ollen mahdollinen vaihtoehto kasviperäisten ravintolisien käytölle, erityisesti sellaisten kasvien osalta, joiden farmakologiset vaikutukset perustuvat useamman kuin yhden yhdisteen yhdistelmään. Tässä minikatsauksessa luetellaan joitakin aiemmin tutkittuja AA-yhdisteitä, joilla on kasvainvastaista aktiivisuutta ja joilla voisi olla tulevaisuutta kliinisessä syöpätutkimuksessa.
Tekijöiden panos
NJ-H osallistui artikkelin ideointiin, kirjoittamiseen ja keskusteluun; CP-P osallistui olennaisesti käsikirjoituksen keskusteluun ja tarkistamiseen; VC-T valmisteli kaikki kuviot ja taulukot; MM-V ja AG-E osallistuivat osaltaan artikkelin kemialliseen osioon; AZ-D osallistui olennaisesti molekyylibiologiseen keskusteluun artikkelista. Kaikki kirjoittajat keskustelivat, tarkistivat ja hyväksyivät julkaistavan käsikirjoituksen lopullisen version.
Rahoitus
Tätä työtä on tukenut Consejo Nacional de Ciencia y Tecnología (CONACYT), Meksiko (apuraha numero 285884). VC-T:tä rahoitti CONACYT (väitöskirja-apurahan numero 267787).
Irintaristiriitoja koskeva lausunto
Tekijät ilmoittavat, että tutkimus tehtiin ilman mitään kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdolliseksi eturistiriidaksi.
Kiitokset
NJ-H kiittää CONACYTia hankkeen rahoittamisesta.
Alonso-Castro, A. J., Villarreal, M. L., Salazar-Olivo, L. A., Gómez-Sánchez, M., Domínguez, F., García-Carrancá, A. (2011). Syövän hoitoon käytetyt meksikolaiset lääkekasvit: farmakologiset, fytokemialliset ja etnobotaaniset tutkimukset. J. Ethnopharmacol. 133, 945-972. doi: 10.1016/j.jep.2010.11.055
PubMed Abstract | CrossRef Full Text | Google Scholar
Arthur, F., Woode, E., Terlabi, E., Larbie, C. (2011). Annona muricata (Linn). vesiuutteen akuutin ja subkroonisen myrkyllisyyden arviointi eläimille. Eur. J. Exp. Biol. 1 (4), 115-124.
Google Scholar
Beck, W. T., Cirtain, M. C., Glover, C. J., Felsted, R. L., Safa, A. R. (1988). Indolialkaloidien vaikutukset monilääkeresistenssiin ja P-glykoproteiinin merkitsemiseen vinblastiinin fotoaffiniteettianalogilla. Biochem. Biophys. Res. Commun. 153, 959-966. doi: 10.1016/S0006-291X(88)81321-4
PubMed Abstract | CrossRef Full Text | Google Scholar
Brindis, F., González-Trujano, M. E., González-Andrade, M., Aguirre-Hernández, E., Villalobos-Molina, R. (2013). Annona macroprophyllatan vesiuute: potentiaalinen α-glukosidaasin estäjä. Biomed Res. Int. 443 (2), 495-499. doi: 10.1155/2013/591313
CrossRef Full Text | Google Scholar
Cafferkey, C., Chau, I. (2016). Uudet STAT3-estäjät mahasyövän hoidossa. Expert Opin. Investig. Drugs 25, 1023-1031. doi: 10.1080/13543784.2016.1195807
PubMed Abstract | CrossRef Full Text | Google Scholar
Chávez, D., Mata, R. (1998). Purpurediolin ja purpureniini, kaksi uutta sytotoksista vierekkäistä bis-tetrahydrofuraani-annonaattiasetogeniinia Annona purpurean siemenistä. J. Nat. Prod. 61, 580-584. doi: 10.1021/np970410
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, Y., Chen, J., Xu, S., Wang, Y., Li, X., Cai, B., et al. (2012a). Annonaattisten asetogeniinien kasvainvastainen aktiivisuus HepS- ja S180-ksenografeja kantavissa hiirissä. Bioorg. Med. Chem. Lett. 22, 2717-2719. doi: 10.1016/j.bmcl.2012.02.109
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, Y., Sh, Xu, Chen, J., Wang, Y., Xu, H., Fan, N., et al. (2012b). Annona squamosa -siemenuutteen kasvainvastainen aktiivisuus, joka sisältää annonaattisia asetogeniiniyhdisteitä. J. Ethnopharmacol. 142, 462-466. doi: 10.1016/j.jep.2012.05.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, Y., Chen, J., Wang, Y., Xu, S., Li, X. (2012c). Kuusi sytotoksista annona-acetogeniinia Annona squamosa-siemenistä. Food Chem. 135, 960-966. doi: 10.1016/j.foodchem.2012.05.041
PubMed Abstract | CrossRef Full Text | Google Scholar
Chen, Y., Chen, Y., Shi, Y., Shi, Y., Ch, Ma, Wang, X., Li, Y., et al. (2016). Annona squamosa -siemenöljyn kasvainvastainen aktiivisuus. J. Ethnopharmacol. 193, 362-367. doi: 10.1016/j.jep.2016.08.036
PubMed Abstract | CrossRef Full Text | Google Scholar
Da Rocha, A. B., Lopes, R. M., Schwartsmann, G. (2001). Luonnontuotteet syövänvastaisessa hoidossa. Curr. Opin. Pharmacol. 1, 364-369. doi: 10.1016/S1471-4892(01)00063-7
PubMed Abstract | CrossRef Full Text | Google Scholar
Elisya, Y., Kardono, L. B. S., Simanjuntak, P. (2014). Tabletin muotoilu soursopin (Annona muricata L). lehtien etyyliasetaattiliukoisesta uutteesta. AJAS 2 (3), 323-329.
Google Scholar
Figueroa-González, G., Jacobo-Herrera, N., Zentella-Dehesa, A., Pereda-Miranda, R. (2012). Monilääkeresistenssin kumoaminen tai monilääkeresistenssi aamunkoiton hartsiglykosideilla ihmisen rintasyöpäsoluissa. J. Nat. Prod. 75 (1), 93-97. doi: 10.1021/np200864m
PubMed Abstract | CrossRef Full Text | Google Scholar
Figueroa-González, G., García-Castillo, V., Coronel-Hernández, J., López-Urrutida, E., León-Cabrera, S., Arias-Romero, L. E., et al. (2016). Koliittiin liittyvän kolorektaalisyövän kolmoishoidon anti-inflammatorinen ja kasvainvastainen aktiivisuus. J. Cancer 7 (12), 1632-1644. doi: 10.7150/jca.13123
PubMed Abstract | CrossRef Full Text | Google Scholar
García-Castillo, V., López-Urrutia, E., Villanueva-Sánchez, O., Ávila-Rodríguez, MÁ, Zentella-Dehesa, A., Cortés-González, C., et al. (2017). Metabolisen uudelleenmuotoilun kohdentaminen kolmoisnegatiivisessa rintasyövässä hiirimallissa. J. Cancer 8 (2), 178-189. doi: 10.7150/jca.16387
PubMed Abstract | CrossRef Full Text | Google Scholar
He, C., Klionsky, D. J. (2009). Autofagian säätelymekanismit ja signaalireitit. Annu. Rev. Genet. 43, 67-93. doi: 10.1146/annurev-genet-102808-114910
PubMed Abstract | CrossRef Full Text | Google Scholar
Igney, F. H., Krammer, P. H. (2002). Kuolema ja kuolemanvastaisuus: kasvainten vastustuskyky apoptoosia vastaan. Nat. Rev. Cancer 2 (4), 277-288. doi: 10.1038/nrc776
PubMed Abstract | CrossRef Full Text | Google Scholar
Jacobo-Herrera, N. J., Jacobo-Herrera, F. E., Zentella-Dehesa, A., Andrade-Cetto, A., Heinrich, M., Pérez-Plasencia, C. (2016). Meksikolaisessa perinteisessä lääketieteessä käytetyt lääkekasvit paksusuolen syövän hoidossa. J. Ethnopharmacol. 179, 391-402. doi: 10.1016/j.jep.2015.12.042
PubMed Abstract | CrossRef Full Text | Google Scholar
Ji, L., Yuan, Y., Luo, L., Chen, Z., Ma, X., Ma, Z., et al. (2012). Physaliinit, joilla on anti-inflammatorinen aktiivisuus, esiintyvät Physalis alkekengi var. franchetii -kasvissa ja voivat toimia Michael-reaktion akseptoreina. Steroids 77, 441-447. doi: 10.1016/j.steroids.2011.11.016
PubMed Abstract | CrossRef Full Text | Google Scholar
Jodoin, J., Demeule, M., Béliveau, R. (2002). Monilääkeresistenssin P-glykoproteiinin aktiivisuuden estäminen vihreän teen polyfenoleilla. Biochim. Biophys. Acta 1542 (1-3), 149-159. doi: 10.1016/S0167-4889(01)00175-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Kenific, C. M., Debnath, J. (2015). Autofagian solu- ja aineenvaihduntatoiminnot syöpäsoluissa. Trends Cell Biol. 25 (1), 37-45. doi: 10.1016/j.tcb.2014.09.001
PubMed Abstract | CrossRef Full Text | Google Scholar
Ko, Y. M., Wu, T. Y., Wu, Y. C. h., Chang, F.r., Guh, J.Y., Chuang, L. Y. (2011). Annonasiini indusoi solusykli-riippuvaisen kasvupysähdyksen ja apoptoosin estrogeenireseptoriin-α liittyvillä reiteillä MCF-7-soluissa. J. Ethnopharmacol. Annona cornifolia A. St.-Hilistä eristettyjen asetogeniinien in vitro sytotoksiset, sienilääkkeelliset, trypanosidiset ja leishmanisidiset vaikutukset. (Annonaceae). An. Acad. Bras. Cienc. 86 (2), 829-839. doi: 10.1590/0001-3765201420130048
PubMed Abstract | CrossRef Full Text | Google Scholar
Liu, Y. Q., Cheng, X., Guo, L. X., Ch, Mao, YJ, Chen, Liu, H. X., et al. (2012). Annonaattisten asetogeniinien mimeettisen aineen, AA05, tunnistaminen AMPK-aktivaattoriksi ja autofagian indusoijaksi paksusuolen syöpäsoluissa. PLoS One 7 (10), e47049-e-47049. doi: 10.1371/journal.pone.0047049
CrossRef Full Text | Google Scholar
Mangal, M., Khan, M. I., Agarwal, S. M. (2016). Asetogeniinit potentiaalisina syöpälääkkeinä. Anticancer Agents Med. Chem. 16, 138-159. doi: 10.2174/1871520615666150629101827
CrossRef Full Text | Google Scholar
Miao, Y. J., Shi, Y. Y., Xu, X. F., Chen, Y., Chen, J. W., Li, X. (2016a). Kolme sytotoksista Annona-acetogeniinia Annona squamosan siemenistä. Phytochem. Lett. 16, 92-96. doi: 10.1016/j.phytol.2016.03.006
CrossRef Full Text | Google Scholar
Miao, Y. J., Shi, Y. Y., Li, F. Q., Shan, C. h. X., Chen, Y., Chen, J. W., et al. (2016b). Metabolomitutkimus Annona squamosan myrkyllisyydestä ultrasuorituskykyisen nestekromatografian ja korkearesoluutioisen massaspektrometrian avulla yhdistettynä hahmontunnistusmenetelmään ja metabolisten polkujen analyysiin. J. Ethnopharmacol. 184, 187-195. doi: 10.1016/j.jep.2016.03.006
PubMed Abstract | CrossRef Full Text | Google Scholar
Moghadamtousi, S. Z., Fadaeinasab, M., Nikzad, S., Mohan, G., Ali, H. M., Kadir, H. A. (2015a). Annona muricata (Annonaceae): katsaus sen perinteiseen käyttöön, eristettyihin asetogeniineihin ja biologisiin vaikutuksiin. Int. J. Mol. Sci. 16, 15625-15658. doi: 10.3390/ijms160715625
PubMed Abstract | CrossRef Full Text | Google Scholar
Moghadamtousi, S. Z., Rouhollahi, E., Karimian, H., Fadaeinasab, M., Firoozinia, M., Abdulla, M. A., et al. (2015b). Annona muricata -lehtien kemopotentiaalinen vaikutus atsoksimetaanin aiheuttamia paksusuolen poikkeavia kryptofokuksia vastaan rotilla ja asetogeeni annomurisiini E:n apoptoosivaikutus HT-29-soluissa: biotestiohjattu lähestymistapa. PLoS One 10 (4), e0122288. doi: 10.1371/journal.pone.0122288
PubMed Abstract | CrossRef Full Text | Google Scholar
Mondal, S. K., Mondal, N. B., Mazumder, U. K. (2007). Annona reticulata -lehtien in vitro -sytotoksinen ja ihmisen rekombinantti kaspaasivaikutus. Indian J. Pharmacol. 39, 253-254. doi: 10.4103/0253-7613.37279
CrossRef Full Text | Google Scholar
Newman, D. J., Cragg, G. M. (2016). Luonnontuotteet uusien lääkkeiden lähteinä vuodesta 1981 vuoteen 2014. J. Nat. Prod. 79 (3), 629-661. doi: 10.1021/acs.jnatprod.5b01055
PubMed Abstract | CrossRef Full Text | Google Scholar
Nobili, S., Lippi, D., Witort, E., Donnini, M., Bausi, L., Mini, E., et al. (2009). Syövän hoidossa ja ennaltaehkäisyssä käytettävät luonnonyhdisteet. Pharmacol. Res. 59, 365-378. doi: 10.1016/j.phrs.2009.01.017
PubMed Abstract | CrossRef Full Text | Google Scholar
OECD (2008). ”Teksti nro 425: Välitön myrkyllisyys suun kautta: ylös ja alas -menettely” teoksessa OECD:n kiltalinjat kemikaalien testausta varten, jakso 4. OECDiLibrary. https://www.oecd-ilibrary.org/environment/test-no-425-acute-oral-toxicity-up-and-down-procedure_9789264071049_en
Google Scholar
Okada, H., Mak, T. W. (2004). Apoptoottisen ja ei-apoptoottisen kuoleman polut kasvainsoluissa. Nat. Rev. Cancer 4 (8), 592-603. doi: 10.1038/nrc1412
PubMed Abstract | CrossRef Full Text | Google Scholar
Pardhasaradhi, B. V., Reddy, M., Ali, A. M., Kumari, A. L., Khar, A. (2005). Annona squamosan siemenuutteiden erilaiset sytotoksiset vaikutukset ihmisen kasvainsolulinjoihin: reaktiivisten happilajien ja glutationin rooli. J. Biosci. 30, 237-244. doi: 10.1007/BF02703704
PubMed Abstract | CrossRef Full Text | Google Scholar
Phang, J. M., Poore, M., Lopaczynska, J., Yeh, G. C. (1993). Flavonolia stimuloiva 7,12-dimetyylibents(a)antraseenin effluksi monilääkeresistenteissä rintasyöpäsoluissa. Cancer Res. 53, 5977-5981.
PubMed Abstract | Google Scholar
Qian, J. Q., Sun, P., Pan, Z. Y., Fang, Z. Z. (2015). Annonaceous-asetogeniinit kääntävät ihmisen hepatosellulaarisen karsinooman BEL-7402/5-FU- ja HepG2/ADM-solulinjojen lääkeresistenssin. Int. J. Clin. Exp. Pathol. 8 (9), 11934-11944.
PubMed Abstract | Google Scholar
Raynaud, S., Némati, F., Miccoli, L., Michel, P., Poupon, M. F., Fourneau, C., et al. (1999). Squamocinin antitumoraaliset vaikutukset alkuperäisiin ja monilääkeresistentteihin MCF7-solulinjoihin (ihmisen rinta-adenokarsinooma). Life Sci. 65 (5), 525-533. doi: 10.1016/S0024-3205(99)00273-8
PubMed Abstract | CrossRef Full Text | Google Scholar
Savithramma, N., Rao, M. L., Suhrulatha, D. (2011). Lääkekasvien seulonta sekundaaristen metaboliittien osalta. MEJSR 8 (3), 579-584.
Google Scholar
Schlie-Guzmán, M. A., García-Carrancá, A., González-Esquinca, A. R. (2009). Annona diversifolia Saff:n laherraduriinin ja cherimolin-2:n in vitro ja in vivo antiproliferatiivinen aktiivisuus. Phytother. Res. 23, 1128-1133. doi: 10.1002/ptr.2760
PubMed Abstract | CrossRef Full Text | Google Scholar
Silva, G. L., Cui, B., Chávez, D., You, M., Hee-Byung, C., Rasoanaivo, P., et al. (2001). Monilääkeresistenssi-fenotyypin modulointi Erythroxylum pervillei -kasvintuhoojasta peräisin olevilla uusilla aromaattisilla trotaanialkaloidiesterillä. J. Nat. Prod. 64 (12), 1514-1520. doi: 10.1021/np010295+
PubMed Abstract | CrossRef Full Text | Google Scholar
Sousa, O. V., Vieira, G. D., Pinho, JJRGd, Yamamoto, C. H., Alves, M. S. (2010). Annona muricata L. -lehtien etanoliuutteen antinociceptiiviset ja anti-inflammatoriset vaikutukset eläinmalleissa. Int. J. Mol. Sci. 11 (5), 2067-2078. doi: 10.3390/ijms11052067
PubMed Abstract | CrossRef Full Text | Google Scholar
Sultana, S., Asif, H. M., Nazar, H. M. I., Akhtar, N., Rehman, J. U., Rehman, R. U. (2014). Syöpää vastaan taistelevat lääkekasvit – vihreä syövänvastainen lähestymistapa. Asian Pac. J. Cancer P 15 (11), 4385-4394. doi: 10.7314/APJCP.2014.15.11.4385
CrossRef Full Text | Google Scholar
Tantithanaporn, S., Wattanapiromsakul, C., Itharat, A., Keawpradub, N. (2011). Goniothalamus undulatus Ridl. -juuriuutteista eristettyjen asetogeniinien ja styrylilaktonien sytotoksinen aktiivisuus keuhkosyöpäsolulinjaa (COR-L23) vastaan. Phytomedicine 18, 486-490. doi: 10.1016/j.phymed.2010.10.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Tormo, J. R., Gallardo, T., Aragón, R., Cortes, D., Estornell, E. (1999). Monotetrahydrofuraanisten annonacetogeniinien spesifiset vuorovaikutukset mitokondriokompleksi I:n estäjinä. Chem. Biol. Interact. 122, 171-183. doi: 10.1016/S0009-2797(99)00120-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Torres, M. P., Rachagani, S., Purohit, V., Pandey, P., Joshi, S., Moore, E. D., et al. (2012). Graviola: uusi lupaava luonnosta peräisin oleva lääkeaine, joka estää haimasyöpäsolujen tuumorigeniteettiä ja metastaasia in vitro ja in vivo muuttamalla solujen aineenvaihduntaa. Cancer Lett. 323 (1), 29-40. doi: 10.1016/j.canlet.2012.03.03.031
PubMed Abstract | CrossRef Full Text | Google Scholar
Wang, L. Q., Min, B. S., Li, Y., Nakamura, N., Qin, G. W., Li, C. J., et al. (2002). Annona montanan lehdistä peräisin olevat annonaattiset asetogeniinit. Bioorg. Med. Chem. 10, 561-565. doi: 10.1016/S0968-0896(01)00303-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Yang, C., Gundala, R., Mukkavilli, R., Vangala, S., Reid, M. D., Aneja, R. (2015). Flavonoidien ja asetogeniinien synergistinen vuorovaikutus Graviolan (Annona muricata) lehdissä antaa suojan eturauhassyöpää vastaan. Carcinogenesis 36 (6), 656-665. doi: 10.1093/carcin/bgv046
PubMed Abstract | CrossRef Full Text | Google Scholar
Yiallouris, A., Patrikios, I., Johnson, E. O., Sereti, E., Dimas, K., De Ford, C., et al. (2018). Annonasiini edistää selektiivistä syöpäsolukuolemaa NKA-riippuvaisten ja SERCA-riippuvaisten reittien kautta. Cell Death Dis. 9 (764), 13. doi: 10.1038/s41419-018-0772-x
PubMed Abstract | CrossRef Full Text | Google Scholar
Yuan, F., Bai, G., Chen, Y., Miao, Y., Chen, J., Li, X. (2015). Erilaisten ACG:iden rakenne-aktiivisuussuhteet monilääkeresistenttiä ihmisen keuhkosyöpäsolulinjaa A549/Taxol vastaan. Bioorg. Med. Chem. Lett. 25, 787-790. doi: 10.1016/j.bmcl.2014.12.088
PubMed Abstract | CrossRef Full Text | Google Scholar
Yuan, S. S., Chang, H. L., Chen, H. W., Chen, H. T. W., Yeh, Y. T., Y. T., Kao, Y. H., Lin, K. H., et al. (2003). Annonasiini, mono-tetrahydrofuraani asetogeniini, pysäyttää syöpäsolut G1-vaiheessa en aiheuttaa sytotoksisuutta Bax. Ja kaspaasi-3:een liittyvää reittiä. Life Sci. 72, 2853-2861. doi: 10.1016/S0024-3205(03)00190-5
PubMed Abstract | CrossRef Full Text | Google Scholar
Yuan, S. S., Chang, H. L., Chen, H. W., Kuo, F. c. h., Liaw, C. h. C. h., Su, J. H., et al. (2006). Squamocinin selektiivinen sytotoksisuus T24-virtsarakon syöpäsoluihin S-vaiheessa Bax-, Bad- ja kaspaasi-3:een liittyvien reittien kautta. Life Sci. 78, 869-874. doi: 10.1016/j.lfs.2005.05.068
PubMed Abstract | CrossRef Full Text | Google Scholar
Zeng, L., Wu, F. E., Oberlies, N. H., McLaughlin, J. L., Sastrodihajo, S. (1996). Viisi uutta monotetrahydrofuraanirenkaan asetogeniinia Annona muricatan lehdistä. J. Nat. Prod. 59, 1035-1042. doi: 10.1021/np960447e
PubMed Abstract | CrossRef Full Text | Google Scholar
Zhou, S., Lim, L. Y., Chowbay, B. (2004). P-glykoproteiinin kasviperäinen modulaatio. Drug Metab. Rev. 36 (1), 57-104. doi: 10.1081/DMR-120028427
PubMed Abstract | CrossRef Full Text | Google Scholar