- Introduction
- MS/DBB Kolinergisten hermosolujen aktiivisuus ja asetyylikoliinin vapautuminen Hippokampuksen CA1:ssä
- Muskariinireseptorin aktivaation vaikutukset hippokampuksen CA1:n inhiboiviin interneuroneihin
- Muskariinireseptorien aktivoituminen Hippokampuksen CA1-interneuroneissa asetyylikoliinin vapautumisen jälkeen
- Nikotiinireseptorin aktivoinnin vaikutukset Hippokampuksen CA1-interneuroneihin
- Nikotiinireseptorien aktivoituminen Hippokampuksen CA1-interneuroneissa asetyylikoliinin vapautumisen jälkeen
- Asetyylikoliinin vapautumisen vaikutukset Hippokampuksen CA1-verkon toimintaan interneuronien membraanipotentiaalin näkökulmasta
- Erittely eturistiriidasta
- Kiitokset
Introduction
Asetyylikoliinia vapautuu koko nisäkkäiden keskushermostossa (CNS), jossa se vaikuttaa aivojen globaaliin toimintaan vaikuttamalla unen- ja heräämisrytmiin, tarkkaavaisuuteen ja muistin muodostumiseen. Yksi aivojen alue, jota hermottavat voimakkaasti mediaalisen septumin ja Brocan kompleksin diagonaalikaistan (MS/DBB) kolinergiset afferentit, on hippokampus (Dutar ym., 1995). Toiminnallisesti on ehdotettu, että asetyylikoliinin vapautuminen hippokampuksessa auttaa muistojen muodostamisessa tai palauttamisessa riippuen solunulkoisesta asetyylikoliinipitoisuudesta (Power et al., 2003; Hasselmo ja Giocomo, 2006; Kenney ja Gould, 2008; Deiana et al., 2011; Hasselmo ja Sarter, 2011; Easton et al., 2012; Blake et al., 2014). Mekanismi, jolla MS/DBB:n kolinergiset päätelaitteet vaikuttavat hippokampusverkon toimintaan, on sekä muskariini- että nikotiinireseptorien aktivoituminen, jotka sijaitsevat pyramidineuronien ja inhibitoristen interneuronien dendriiteissä, solurungoissa ja aksoniterminaaleissa sekä astrosyyttien päätelaitteissa (Cobb ja Davies, 2005; Teles-Grilo Ruivo ja Mellor, 2013). Vaikka asetyylikoliini vaikuttaa useisiin paikkoihin useissa eri solutyypeissä, osa sen vaikutuksesta syntyy todennäköisesti sen vaikutuksista interneuronien toimintaan.
Inhiboivilla interneuroneilla on ratkaiseva rooli tiedonkäsittelyssä hippokampuksessa. Interneuronit ovat anatomiselta rakenteeltaan ja oletetulta toiminnaltaan hyvin erilaisia (Freund ja Buzsaki, 1996; Klausberger ja Somogyi, 2008). Riippuen interneuronin alatyypistä ja siitä, missä se innervoi pyramidisolua, yksittäinen interneuroni voi kokonaan estää aktiivisuuden dendriitissä, muuttaa toimintapotentiaalin laukaisuvaihetta somen kohdalla tai estää kokonaan toimintapotentiaalin laukaisun pyramidisolun rungossa (Miles et al., 1996; Larkum et al., 1999). Verkkotasolla interneuronit edistävät synkronisen aktiivisuuden syntymistä pääneuronien populaatioiden välillä erilaisilla käyttäytymisen kannalta merkityksellisillä taajuuksilla (Buzsaki, 2002; Buzsaki ja Wang, 2012). Koska yksittäisillä interneuroneilla on merkittävä vaikutus neuroniverkon toimintaan, on todennäköistä, että huomattava osa asetyylikoliinin vaikutuksesta hippokampuksen aktiivisuuteen syntyy interneuronien moduloinnin kautta. Vaikka kolinergisten reseptorien on osoitettu vaikuttavan inhiboiviin presynaptisiin päätelaitteisiin (Behrends ja Ten Bruggencate, 1993; Tang ym., 2011) ja interneuronien herätettävyyteen (McQuiston ja Madison, 1999b; Griguoli ym., 2009; Cea-Del Rio ym, 2010, 2011), tässä minikatsauksessa keskitytään viimeaikaisiin tutkimuksiin, joissa on tutkittu asetyylikoliinin vapautumisen vaikutusta interneuronien kalvopotentiaalin muutoksiin, erityisesti hippokampuksen CA1:ssä.
MS/DBB Kolinergisten hermosolujen aktiivisuus ja asetyylikoliinin vapautuminen Hippokampuksen CA1:ssä
Asetyylikoliinin vapautumisen vaikutus hippokampuksen CA1:ssä ja se, missä määrin se vaikuttaa eri interneuronien alatyyppeihin, riippuu kolinergisten aksonien päätelaitteiden spesifisestä sijainnista ja tiheydestä sekä sen inaktivoivasta entsyymistä, asetyylikoliiniesteraasista. Huomattavaa on, että sekä kolinergisten kuitujen että asetyylikoliiniesteraasin on osoitettu jakautuvan eri tavoin eri kerroksiin hippokampuksen CA1:ssä. Hiirellä kolinergisten kuitujen osoitettiin jakautuvan tasaisesti lukuun ottamatta kahta tiheämpää kaistaa stratum pyamidalessa (SP) ja stratum radiatumin (SR) ja stratum lacunosum-molecularen (SLM) rajalla (Aznavour et al., 2002). Rotilla samanlaisia tiheämpiä kaistoja havaittiin SP:ssä sekä SR:n ja SLM:n rajalla. Verrattuna stratum oriensiin (SO) SR:ssä havaittiin kuitenkin pienempiä tiheyksiä ja SLM:ssä vielä pienempiä tiheyksiä (SO > SR > SLM) (Aznavour et al., 2002). Asetyylikoliiniesteraasin jakautuminen hippokampuksen CA1:ssä täydentää kolinergisen syötteen jakautumista, ja korkeampia tiheyksiä havaittiin SP:n ja SO:n välillä sekä toinen huippu SLM:ssä lähellä SR:n rajaa (Storm-Mathisen, 1970). Yhdenmukaisesti näiden anatomisten tietojen kanssa asetyylikoliinin lisääntyneen vapautumisen mittaukset theta-rytmien aikana ovat osoittaneet, että asetyylikoliinipitoisuudet olivat korkeimmat lähellä stratum pyramidalea (Zhang et ai., 2010). Tämä kolinergisten kuitujen ja solunulkoisten asetyylikoliinipitoisuuksien erilainen jakautuminen on erityisen tärkeää, kun otetaan huomioon, että kaikki kolinergiset terminaalit hippokampuksessa eivät näytä välittävän asetyylikoliinia synaptisesti. Sekä hippokampuksessa että neokorteksissa arvioitiin, että 85-93 prosentilla kolinergisistä aksoniterminaaleista ei ole postsynaptista erikoistumista, ja näin ollen suurimmalla osalla kolinergisistä terminaaleista ehdotettiin välitettävän asetyylikoliinia volyymin tai ei-synaptisen välityksen avulla (Umbriaco et al., 1994, 1995). Toiset ryhmät ovat kuitenkin arvioineet, että suurin osa kolinergisistä päätepisteistä (66-67 %) neokorteksissa muodostaa klassisia synaptisia yhteyksiä (Smiley ym., 1997; Turrini ym., 2001). Tästä ristiriidasta huolimatta merkittävä osa pääteistä näyttää vapauttavan asetyylikoliinia solunulkoiseen tilaan parakriinin kaltaisella tavalla. Tämä edellyttää, että terminaalisesti vapautuva asetyylikoliini diffundoituu merkittäviä matkoja asetyylikoliiniesteraasin ohi sitoutuakseen postsynaptisten elementtien reseptoreihin. Näin ollen alueet tai kerrokset, joilla on suotuisia tiheyksiä kolinergisiä päätelaitteita (korkeampia) ja/tai asetyylikoliiniesteraasia (matalampia), voivat johtaa suurempiin solunulkoisiin asetyylikoliinipitoisuuksiin, jotka voivat olla tehokkaampia välittämään asetyylikoliinia tilavuussiirron kautta. Lisäksi on mahdollista, että on olemassa osajoukko päätelaitteita, jotka ovat aktiivisempia, joiden vapautumisen todennäköisyys on suurempi tai jotka voivat vapauttaa enemmän välittäjäainetta. Nämä päätelaitteet voivat olla tehokkaampia välittämään tilavuussiirtoa ja vaikuttamaan läheisiin inhiboiviin interneuroneihin.
Asetyylikoliinin vapautuminen kolinergisistä päätelaitteista riippuu MS/DBB:n kolinergisten neuronien aktiivisuudesta. Kirjallisuudessa raportoidut MS/DBB:n kolinergisten neuronien laukaisumallit ovat kuitenkin osoittaneet jonkin verran vaihtelua (Barrenechea et al., 1995; Brazhnik ja Fox, 1997, 1999; Simon et al., 2006). Pienellä määrällä anatomisesti tunnistettuja MS/DBB-kolinergisiä neuroneja, jotka on nauhoitettu hereillä olevilla hillityillä jyrsijöillä, on raportoitu olevan alhainen epäsäännöllinen ampumisnopeus (<2 Hz) (Simon et ai., 2006). Sitä vastoin anatomisesti tunnistamattomien neuronien, joiden toimintapotentiaalien aaltomuodot ovat yhdenmukaisia MS/DBB-kolinergisten neuronien kanssa, on raportoitu laukeavan jopa 30 Hz:n taajuudella (Brazhnik ja Fox, 1999). Näin ollen on edelleen epäselvää, mitkä nopeudet kuvaavat parhaiten MS/DBB:n kolinergisten neuronien laukaisukuvioita vai kuuluvatko ne laajaan jatkumoon. Mahdollisilla eroilla kolinergisten neuronien laukaisutaajuudessa tai aktiivisuuden kestossa voisi kuitenkin olla vaihtelevia vaikutuksia eri interneuronien alatyyppeihin asetyylikoliinipitoisuuksien paikallisten erojen kautta.
Muskariinireseptorin aktivaation vaikutukset hippokampuksen CA1:n inhiboiviin interneuroneihin
Muskariinireseptorien systeemisellä estolla tai muskariinireseptoriantagonistien suoralla injektoinnilla hippokampukseen tapahtuva MS/DBB:n kolinergeettisen toiminnan häirintä voi heikentää muistia ja spatiaalisen informaation koodausta (Blokland ym, 1992; Atri ym., 2004; Hasselmo, 2006). Inhibitoristen interneuronien mahdollinen rooli hippokampuksen toiminnan muskariinireseptorimodulaatiossa perustui alun perin havaintoihin, joiden mukaan kolinergisten agonistien eksogeeninen käyttö johti spontaanien inhibitoristen postsynaptisten virtojen (sIPSC) lisääntymiseen CA1-pyramidineuroneissa (Pitler ja Alger, 1992). Nämä tiedot viittasivat epäsuorasti siihen, että osa inhiboivista interneuroneista voi depolarisoitua muskariinireseptorin aktivaatiolla, ja ne vahvistettiin myöhemmin suorilla rekisteröinneillä (Parra et al., 1998; McQuiston ja Madison, 1999a). Kaikki interneuronit eivät kuitenkaan vastanneet muskariinireseptorin aktivoitumiseen depolarisaatiolla. Jotkut interneuronit hyperpolarisoituivat tai osoittivat kaksivaiheisia vasteita, ja jotkut eivät vastanneet muskariinisen agonistin eksogeeniseen annosteluun (Parra et al., 1998; McQuiston ja Madison, 1999a). Lisäksi kutakin muskariinivasteen tyyppiä ei voitu korreloida interneuronin morfologisen alatyypin kanssa. Näitä havaintoja vaikeutti entisestään havainto, että muskariinireseptorit voivat estää GABA:n vapautumista perisomaattisten inhibitoristen interneuronien osajoukosta (Behrends ja Ten Bruggencate, 1993; Fukudome ym., 2004; Szabo ym., 2010) ja muskariinireseptorin aktivaatio voi lisätä interneuronien eksitoituvuutta synnyttämällä jälkidepolarisaatioita (McQuiston ja Madison, 1999b; Lawrence ym., 2006). Näin ollen asetyylikoliinin vapautumisen vaikutus interneuronipopulaatioon on monimutkainen ja johtaa joidenkin interneuronien rekrytointiin samalla kun se estää toisia.
Muskariinireseptorien aktivoituminen Hippokampuksen CA1-interneuroneissa asetyylikoliinin vapautumisen jälkeen
Vaikka kolinergiset muskariiniset synaptiset vasteet mitattiin ensimmäisen kerran CA1-pyramidineuroneissa vuonna 1983 (Cole ja Nicoll, 1983), vasta vuonna 2006 mitattiin muskariiniset vasteet sähköisesti herätettyyn asetyylikoliinin vapautumiseen Hippokampuksen CA1-estäjävälitteisissä interneuroneissa (Widmer et al., 2006). Tämä tutkimus osoitti, että terminaalisesti vapautuvalla asetyylikoliinilla oli erilaiset vaikutukset eri interneuronien alatyyppeihin. Interneuronit saattoivat reagoida depolarisoimalla, hyperpolarisoimalla tai kaksivaiheisilla vasteilla. Kaiken kaikkiaan suurin osa reagoivista interneuroneista tuotti depolarisaatioita (64 %), kun taas hyperpolarisaatioita havaittiin harvoin (13 %) (Widmer ym., 2006). Lisäksi, kuten aiemmissa tutkimuksissa, joissa käytettiin muskariinisten agonistien eksogeenista käyttöä (Parra et al., 1998; McQuiston ja Madison, 1999a), erilaisia sähköisesti herätettyjä muskariinisia vastetyyppejä ei voitu korreloida tiettyjen interneuronien anatomisten alatyyppien kanssa (Widmer et al., 2006). Nämä havainnot on äskettäin vahvistettu optogeneettisissä tutkimuksissa, joissa käytettiin evokoitua vapautumista vasteena valoaktivointiin (Nagode et ai., 2011; Bell et ai., 2013). Yhdessä näistä optogeneettisistä tutkimuksista interneuronit, jotka vastasivat bifaasisilla (25 %), hyperpolarisoivilla (35 %) ja depolarisoivilla (40 %) muskariinivasteilla, jakautuivat kuitenkin tasaisemmin eri vastetyyppien kesken (Bell et ai., 2013). Tärkeää on, että optogeneettisesti vapautettu asetyylikoliini tuotti pääasiassa muskariinivasteet (80%) vs. nikotiinivasteet (17%). Loput 3 % vastanneista interneuroneista sai sekä muskariinisia että nikotiinisia vasteita. Lisäksi muskariiniset hyperpolarisaatiot välitettiin M4-reseptorien aktivoinnilla, kun taas depolarisaatiot tuotettiin todennäköisesti M3-reseptorien aktivoinnilla (Bell ym., 2013). Sähköstimulaatiotutkimusten tapaan muskariinivasteen tyyppiä ei voitu korreloida anatomisten interneuronien alatyyppien kanssa. Tärkeää on, että molemmat tutkimukset osoittivat, että perisomaattisesti projisoivat interneuronit (todennäköisesti parvalbumiinia ilmentävät korisolut) voivat reagoida asetyylikoliinin vapautumiseen millä tahansa kolmesta muskariinisesta vastetyypistä (Widmer et al., 2006; Bell et al., 2013). Erilaisissa optogeneettisissä tutkimuksissa CA1-interneuronien kalvopotentiaalia arvioitiin epäsuorasti mittaamalla sIPSC-taajuutta CA1-pyramidineuroneissa (Nagode et al., 2011, 2014). Optogeneettisesti vapautettu asetyylikoliini johti suurten amplitudisten sIPSC:ien lisääntymiseen taajuuksilla, jotka kuuluivat theta-kaistanleveyteen (4-12 Hz) (Nagode et al., 2011). Tärkeää on, että endokannabinoidit pystyivät estämään tämän sIPSC:n lisääntymisen, mikä viittaa siihen, että ne johtuivat kolekystokiniinipositiivisten interneuronien aktivoitumisesta (Nagode et al., 2011). Lisäksi parvalbumiinipositiivisten solujen optogeneettinen tukahduttaminen ei vaikuttanut sIPSC:iin, mikä viittaa siihen, että ne eivät johtuneet parvalbumiinikorisolujen, akso-aksonisten, bistratifioitujen tai oriens-lacunosum-moleculare-interneuronien aktivoitumisesta (Nagode et al., 2014). Nämä havainnot ovat johdonmukaisia synaptisten stimulaatiotutkimusten kanssa, joissa rekisteröitiin kolekystokiniinikorisolujen morfologiaa omaavasta interneuronista, joka tuotti kaksivaiheisen vasteen asetyylikoliinin vapautumiseen (Widmer et ai., 2006). Siksi pelkästään kalvopotentiaaliin kohdistuvien vaikutusten perusteella hippokampuksen CA1- interneuronien endogeenisesti aktivoituneilla muskariinireseptoreilla on monimutkaisia vaikutuksia verkon toimintaan (ks. taulukko 1).
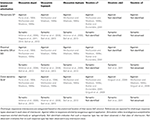
TAULUKKO 1. Kolinergiset vasteet vaihtelevat samankaltaisissa ja erilaisissa anatomisissa interneuronien alatyypeissä.
Vaikka erilaisia muskariinivasteiden tyyppejä havaittiin lähes yhdenmukaisesti CA1- interneuroneissa, kaikkia vastetyyppejä ei voitu yhtä helposti herättää optogeneettisellä stimulaatiolla (Bell et al., 2013). Yhdenmukaisesti joidenkin in vivo -rekisteröintien kanssa (Brazhnik ja Fox, 1999), 20 Hz: n taajuudella annetuilla sinisen valon välähdyksillä vapautuva asetyylikoliini, joka vapautui MS / DBB-kolinergisistä päätelaitteista, pystyi tuottamaan jokaisen vastetyypin hippokampuksen CA1- interneuroneissa (Bell et ai., 2013). Välähdysten määrä vaikutti kuitenkin tietyn vastetyypin havaitsemisen todennäköisyyteen. Hyperpolarisoivissa interneuroneissa 10 välähdystä riitti (91 % hyperpolarisoivista interneuroneista) vasteen havaitsemiseen. Sitä vastoin 10 välähdystä ei riittänyt tuottamaan vastetta suurimmalla osalla depolarisoivista interneuroneista (58 %). Samoin depolarisaatiovaihetta ei voitu havaita suurimmassa osassa bifaasisia interneuroneja (55 %), kun annettiin vain 10 ärsykettä. Siksi muskariiniset hyperpolarisaatiot saattavat vaatia vähemmän presynaptista MS/DBB-kolinergistä aktiivisuutta verrattuna depolarisoiviin vasteisiin hippokampuksen CA1-interneuroneissa. Saattaa olla, että interneuronien herätettävyyden tukahduttaminen on vallitseva vaikutus vasteena alhaisille MS/DBB-kolinergisen aktiivisuuden tasoille.
Nikotiinireseptorin aktivoinnin vaikutukset Hippokampuksen CA1-interneuroneihin
Nikotiinireseptorien aktivoinnilla hippokampuksessa on merkittävä vaikutus fysiologiseen ja patofysiologiseen muistinmuodostukseen (Levin, 2002; Levin et al., 2002, 2009; Buccafusco et al., 2005; Davis ja Gould, 2006, 2009; Nott ja Levin, 2006; Davis et al., 2007). Nisäkkäiden keskushermostossa esiintyvistä 11 eri nikotiinireseptorin alayksiköstä 9:n on raportoitu ilmentyvän hippokampuksen CA1-neuroneissa (Sudweeks ja Yakel, 2000). Käyttämällä nikotiinisten agonistien eksogeenista annostelua funktionaaliset nikotiinireseptorit, jotka sisältävät α7 (Alkondon ym., 1997; Jones ja Yakel, 1997; Frazier ym, 1998b; McQuiston ja Madison, 1999c), α4β2- (McQuiston ja Madison, 1999c; Sudweeks ja Yakel, 2000) tai α2-alayksiköitä (McQuiston ja Madison, 1999c; Sudweeks ja Yakel, 2000; Jia ym., 2009) on havaittu hippokampuksen CA1- interneuroneissa. Vaikka hippokampuksen interneuronit näyttivät ilmentävän erilaisia nikotiinireseptorien alatyyppejä, α7-reseptoreita sisältäviä reseptoreita havaittiin useammin ja ne tuottivat suurempia vasteita (McQuiston ja Madison, 1999c; Sudweeks ja Yakel, 2000). Hippokampuksen α7-nikotiinireseptorit onkin yhdistetty muistinmuodostukseen (Levin, 2002; Levin ym., 2002; Nott ja Levin, 2006), ja niiden toimintahäiriöillä voi olla merkitystä joissakin skitsofrenian muodoissa (Freedman ym., 1994; Leonard ym., 1996; Adler ym., 1998). Alhaisemmasta ilmentymistasostaan huolimatta α4β2-pitoisilla nikotiinireseptoreilla on kuitenkin raportoitu olevan merkittävä rooli muistinmuodostuksessa (Davis ja Gould, 2006; Davis ym., 2007) ja hippokampuksesta riippuvaisessa nikotiiniriippuvuudessa (Perry ym., 2007), 1999; Davis ja Gould, 2009). α4β2-pitoisia reseptoreita on myös korreloitu ikääntymiseen ja Alzheimerin tautiin liittyvien kognitiivisten puutteiden kanssa (Kellar ym., 1987; Wu ym., 2004; Gahring ym., 2005). Jotta eri nikotiinisten alayksiköiden roolia hippokampuksessa voitaisiin täysin ymmärtää, on alettu tutkia endogeenisesti vapautuvan asetyylikoliinin vaikutusta yksittäisiin hippokampussoluihin ja hippokampusverkostoon.
Nikotiinireseptorien aktivoituminen Hippokampuksen CA1-interneuroneissa asetyylikoliinin vapautumisen jälkeen
Asetyylikoliinin vapautumisen MS/DBB-kolinergisistä päätelaitteista hippokampuksen CA1:ssä on osoitettu aktivoivan nikotiinireseptoreita interneuroneissa (Alkondon ym, 1998; Frazier et al., 1998a; Stone, 2007). Nikotiinisia eksitatorisia postsynaptisia virtoja (EPSC) havaittiin ensimmäisen kerran käyttämällä sähköistä stimulaatiota ja koko solun patch clampingia akuuteissa rotan aivoviipaleissa. Näillä nikotiinisilla EPSC:illä oli nopea kinetiikka, ja α7-nikotiinireseptoriantagonistit estivät ne (Alkondon et al., 1998; Frazier et al., 1998a), mikä on yhdenmukaista niiden tutkimusten kanssa, joissa nikotiinireseptoriagonisteja levitettiin suoraan interneuronien soluelimiin (Alkondon et al., 1997; Jones ja Yakel, 1997; Frazier et al., 1998b; McQuiston ja Madison, 1999c). Uudemmissa optogeneettisissä tutkimuksissa hiiren aivoviipaleissa ei kuitenkaan pystytty toistamaan näitä aiempia havaintoja (Bell et al., 2011). Sen sijaan optogeneettisesti vapautettu asetyylikoliini aktivoi ensisijaisesti nikotiinireseptoreita, jotka sisälsivät α4β2-alayksiköitä. Lisäksi α4β2-vasteet olivat enimmäkseen kynnyksen alapuolella ja niillä oli hyvin hidas kinetiikka. Nämä tiedot viittasivat siihen, että asetyylikoliini diffundoi merkittävän matkan ennen sitoutumista α4β2-pitoisiin nikotiinireseptoreihin (McQuiston ja Madison, 1999c; Bennett et ai., 2012), mikä on yhdenmukaista volyymin tai ei-synaptisen siirron kanssa (Vizi et ai., 2010). Vaikka nämä pienet nikotiiniset vasteet voisivat ajallisesti summautua, niiden kyky herättää interneuroneja oli rajoitettu muskariinisen presynaptisen eston kautta. Koska nikotiiniset vasteet olivat enimmäkseen kynnyksen alapuolella, nikotiininen siirto CA1-interneuroneihin voi olla luonteeltaan ensisijaisesti moduloivaa. Optogeneettisissä tutkimuksissa tutkittiin nikotiinisia vasteita myös jänniteherkän väriaineen (VSD) kuvantamisen avulla. Nikotiiniset VSD-signaalit estettiin kokonaan α4β2-reseptoriantagonistilla DHβE, ja niiden havaittiin olevan huomattavasti suurempia CA1-pyramidineuronien distaalisella dendriittisellä alueella, joka on päällekkäinen entorinaalisen aivokuoren ja talamuksen nucleus reuniens -ytimen sisääntulojen kanssa (Bell et al., 2011). Tärkeää on, että koska VSD värjää kaikki kudoksen elementit, VSD-tiedot viittaavat siihen, että α4β2-pitoiset nikotiinireseptorit ovat yleisin reseptori, joka välittää depolarisoivia nikotiinisia vasteita hiiren hippokampuksen CA1:ssä. Huomionarvoista on, että nikotiiniset vasteet voidaan tuottaa yhdellä valon välähdyksellä (Bell et al., 2011), mikä viittaa siihen, että asetyylikoliinin vapautuminen MS/DBB-kolinergisistä päätelaitteista voi auttaa rekrytoimaan interneuroneja nikotiinireseptorin aktivoinnin kautta ennen kuin muskariinireseptorin aktivointi vaikuttaa niihin.
Asetyylikoliinin vapautumisen vaikutukset Hippokampuksen CA1-verkon toimintaan interneuronien membraanipotentiaalin näkökulmasta
Koska CA1:n inhiboivien interneuronien membraanipotentiaalit voivat olla eri tavoin moduloituja sekä muskariini- että nikotiinireseptorin aktivaatiolla asetyylikoliinin vapautumisen jälkeen, siitä johtuva vaikutus verkon toimintaan on epäilemättä monimutkainen. Muskariinireseptorin aktivaatio voi johtaa vaihteleviin ja vastakkaisiin vaikutuksiin jopa saman interneuronin sisällä (ks. taulukko 1). Valitettavasti ymmärryksemme siitä, miten muskariini- tai nikotiinireseptorin aktivaatio voi vaikuttaa kuhunkin interneuronin alatyyppiin, on edelleen puutteellinen. Kunkin vastetyypin tuottamiseen tarvittavien ärsykkeiden määrä vaihteli kuitenkin johdonmukaisesti. Nikotiiniset vasteet olivat helpoimmin herätettävissä, mikä vaati vähiten ärsykkeitä (Bell et al., 2011), kun taas depolarisoivia muskariinisia vasteita oli vaikein tuottaa, mikä vaati eniten ärsykkeitä (Bell et al., 2013). Siksi voidaan olettaa, että alhaiset MS / DBB-kolinergisten neuronien aktiivisuuden tasot ja alhaisemmat solunulkoisen asetyylikoliinin pitoisuudet suosivat nikotiinireseptorien aktivoitumista tai muskariinista hyperpolarisaatiota tietyissä CA1-interneuronien osajoukoissa.
Koska CA1-interneuronien muskariininen hyperpolarisaatio vaatii vähemmän presynaptista kolinergistä aktiivisuutta, hippokampuksen CA1-pyramidisolujen disinhibitiota (epäsuoraa aktivaatiota) voidaan suosia alhaisen MS/DBB-kolinergisen aktiivisuuden aikana (kuva 1B). Lisäksi, jos oletetaan, että nikotiiniset vasteet vaikuttavat ensisijaisesti interneuroneihin, jotka estävät selektiivisesti muita interneuroneja (interneuroni-selektiivinen tai IS), nikotiinireseptorin aktivaatio voi myös johtaa CA1-pyramidineuronien disinhiboitumiseen (kuva 1A). Yhdessä alhaiset MS/DBB-kolinergisen aktiivisuuden tasot edistäisivät hippokampuksen CA1:n nettomääräistä disinhibitiota, mikä mahdollistaisi CA1-pyramidineuronien ulostulon suuremman todennäköisyyden. CA1:n lisääntynyt lähtö voi johtaa muistin palauttamisen ja muistin vakiinnuttamisen helpottumiseen muilla keskushermoston alueilla, kuten ajatellaan tapahtuvan hitaan aallon unen aikana (Gais ja Born, 2004; Hasselmo ja McGaughy, 2004). Sitä vastoin korkeammat MS/DBB-kolinergisten neuronien aktiivisuuden tasot, jotka on yhdistetty korkeampiin solunulkoisiin asetyylikoliinipitoisuuksiin, rekrytoivat myöhemmin erilaisia interneuronien alaryhmiä, jotka reagoivat muskariinisten depolarisaatioiden kautta. Jotkut näistä depolarisoivista interneuroneista voivat määrätä CA1-pyramidineuronien rytmisen eston theta-taajuuksilla (Nagode et ai., 2011, 2014), verkkorytmi, joka havaitaan korkeampien asetyylikoliinin vapautumistasojen aikana (Zhang et ai., 2010). Tämä johtaisi hippokampuksen CA1-pyramidineuronien ulostulon estämiseen (osittain rytminen) ja helpottaisi samalla synaptista integraatiota hippokampuksen CA1-pyramidisolujen dendriittien sisällä kolinergisten vaikutusten kautta glutamatergisiin reseptoreihin ja dendriittiseen toimintaan (kuva 1C) (Tsubokawa ja Ross, 1997; Tsubokawa, 2000; Fernandez De Sevilla ja Buno, 2010; Giessel ja Sabatini, 2010). Tällaista asetyylikoliinipitoisuuksien dynaamista roolia oppimisessa ja muistin muodostumisessa onkin ehdotettu aiemmin (Hasselmo, 2006; Hasselmo ja Giocomo, 2006; Giocomo ja Hasselmo, 2007; Hasselmo ja Sarter, 2011). Tässä järjestelmässä alhaisemmat asetyylikoliinipitoisuudet sallivat hippokampuksen sisäisten (Schafferin kollateraalien) synaptisten vuorovaikutusten hallitsevan, mikä lisää hippokampuksen CA1-tulostusta ja muistin hakua, kun taas korkeammat asetyylikoliinipitoisuudet suosivat hippokampuksen ulkopuolelta tulevien syötteiden käsittelyä, mikä mahdollistaa muistojen ohimenevän muodostumisen hippokampuksen CA1:ssä. Siksi asetyylikoliinin vapautumisen yhteisvaikutus glutamatergisiin tuloihin ja interneuronien toimintaan voi olla tärkeässä roolissa viritettäessä hippokampuksen CA1-verkkoa muistin palauttamista tai uusien muistojen muodostamista varten.

Kuva 1. Hypoteesi, jonka mukaan MS/DBB-kolinergiset tulot joko tukahduttavat tai aktivoivat interneuroniverkostoja hippokampuksen CA1:ssä riippuen kolinergisen neuronin aktiivisuudesta. (A) Alhainen MS/DBB-kolinerginen aktiivisuus aktivoi ensisijaisesti interneuronien osajoukkoja nikotiinireseptorien aktivoinnin kautta. Oletamme, että nikotiini-ohjatut interneuronit ovat interneuronien selektiivisiä interneuroneja (IS, keltainen-aktivointi), jotka estävät spesifisesti muita interneuroneja (sininen). Niiden aktiivisuuden lisääminen johtaa pyramidineuronien (P, keltainen aktivoituminen ja lisääntynyt tuotos) disinhiboitumiseen. (B) Matalat MS/DBB-kolinergisen aktiivisuuden tasot hyperpolarisoivat myös interneuronien osajoukkoja aktivoimalla muskariinireseptoreita (I, sininen-suppressio), mikä johtaa pyramidineuronien disinhiboitumiseen (P, keltainen-aktivoituminen ja lisääntynyt tuotanto). (C) Kolinergisten neuronien aktiivisuuden lisääntyminen aiheuttaa interneuronien osajoukkojen depolarisaation muskariinireseptorien aktivoitumisen kautta (I, punainen-aktivoituminen ja lisääntynyt synaptinen inhibitio), mikä johtaa pyramidineuronien suppressioon (P, sininen-suppressoitu ulostulo).
Erittely eturistiriidasta
Tekijä ilmoittaa, että tutkimus tehtiin ilman kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdolliseksi eturistiriidaksi.
Kiitokset
Tätä työtä on tuettu NIMH/NIH:n 5R01MH094626 ja 1R21MH103695 apurahoilla.
Adler, L. E., Olincy, A., Waldo, M., Harris, J. G., Griffith, J., Stevens, K., et al. (1998). Skitsofrenia, sensorinen porttaaminen ja nikotiinireseptorit. Schizophr. Bull. 24, 189-202. doi: 10.1093/oxfordjournals.schbul.a033320
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Alkondon, M., Pereira, E. F., ja Albuquerque, E. X. (1998). alfa-bungarotoksiini- ja metyylikakonitiini-herkät nikotiinireseptorit välittävät nopeaa synaptista siirtoa rotan hippokampuksen viipaleiden interneuroneissa. Brain Res. 810, 257-263. doi: 10.1016/S0006-8993(98)00880-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Alkondon, M., Pereira, E. F., Barbosa, C. T., and Albuquerque, E. X. (1997). Neuronaalisen nikotiinisen asetyylikoliinireseptorin aktivaatio moduloi gamma-aminovoihapon vapautumista rotan hippokampusviipaleiden CA1-neuroneista. J. Pharmacol. Exp. Ther. 283, 1396-1411.
Pubmed Abstract | Pubmed Full Text
Atri, A., Sherman, S., Norman, K. A., Kirchhoff, B. A., Nicolas, M. M., Greicius, M. D., et al. (2004). Keskeisten kolinergisten reseptorien esto heikentää uuden oppimista ja lisää proaktiivista häirintää sanapari-assosiaatiomuistitehtävässä. Behav. Neurosci. 118, 223-236. doi: 10.1037/0735-7044.118.1.223
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Aznavour, N., Mechawar, N., and Descarries, L. (2002). Aikuisen hiiren ja rotan dorsaalisen hippokampuksen kolinergisen innervaation vertaileva analyysi: kvantitatiivinen immunosytokemiallinen tutkimus. Hippocampus 12, 206-217. doi: 10.1002/hipo.1108
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Barrenechea, C., Pedemonte, M., Nunez, A., and Garcia-Austt, E. (1995). Brocan hermosolujen mediaalisen septikaistan ja diagonaalikaistan in vivo-sisäiset rekisteröinnit: suhteet theta-rytmiin. Exp. Brain Res. 103, 31-40. doi: 10.1007/BF00241962
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Behrends, J. C., ja Ten Bruggencate, G. (1993). Synaptisen eston kolinerginen modulaatio marsun hippokampuksessa in vitro: GABAergisten interneuronien eksitaatio ja GABA-vapautumisen esto. J. Neurophysiol. 69, 626-629.
Pubmed Abstract | Pubmed Full Text
Bell, K. A., Shim, H., Chen, C. K., and McQuiston, A. R. (2011). Hippokampuksen CA1 interneuronien nikotiiniset eksitatoriset postsynaptiset potentiaalit välittyvät pääasiassa nikotiinireseptoreilla, jotka sisältävät alfa4- ja beeta2-alayksiköitä. Neuropharmacology 61, 1379-1388. doi: 10.1016/j.neuropharm.2011.08.024
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Bell, L. A., Bell, K. A., and McQuiston, A. R. (2013). Synaptiset muskariiniset vastetyypit hippokampuksen CA1 interneuroneissa riippuvat erilaisista presynaptisen aktiivisuuden tasoista ja erilaisista muskariinireseptorin alatyypeistä. Neuropharmacology 73, 160-173. doi: 10.1016/j.neuropharm.2013.05.026
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Bennett, C., Arroyo, S., Berns, D., ja Hestrin, S. (2012). Mekanismit, jotka tuottavat kaksikomponenttisia nikotiinisia EPSC: tä kortikaalisissa interneuroneissa. J. Neurosci. 32, 17287-17296. doi: 10.1523/JNEUROSCI.3565-12.2012
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Blake, M. G., Krawczyk, M. C., Baratti, C. M., and Boccia, M. M. (2014). Muistin konsolidoinnin ja uudelleenkonsolidoinnin neurofarmakologia: näkemyksiä keskeisistä kolinergisistä mekanismeista. J. Physiol. Paris. doi: 10.1016/j.jphysparis.2014.04.005.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Blokland, A., Honig, W., ja Raaijmakers, W. G. (1992). Hippokampuksen sisäisten skopolamiini-injektioiden vaikutukset toistuvassa avaruudellisessa hankintatehtävässä rotalla. Psychopharmacology (Berl). 109, 373-376. doi: 10.1007/BF02245886
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Brazhnik, E. S., ja Fox, S. E. (1997). Solunsisäiset rekisteröinnit mediaalisen septin neuroneista hippokampuksen theta-rytmin aikana. Exp. Brain Res. 114, 442-453. doi: 10.1007/PL00005653
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Brazhnik, E. S., ja Fox, S. E. (1999). Toimintapotentiaalit ja suhteet mediaalisen septumin neuronien theta-rytmiin in vivo. Exp. Brain Res. 127, 244-258. doi: 10.1007/s002210050794
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Buccafusco, J. J., Letchworth, S. R., Bencherif, M. ja Lippiello, P. M. (2005). Pitkäkestoinen kognitiivinen paraneminen nikotiinireseptoriagonisteilla: farmakokineettis-farmakodynaamisen epäsuhdan mekanismit. Trends Pharmacol. Sci. 26, 352-360. doi: 10.1016/j.tips.2005.05.007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Buhler, A. V., and Dunwiddie, T. V. (2001). Hippokampuksen stratum oriens interneuronien aktiivisuuden säätely alfa7-nikotiinisilla asetyylikoliinireseptoreilla. Neuroscience 106, 55-67. doi: 10.1016/S0306-4522(01)00257-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Buzsaki, G. (2002). Teeta-oskillaatiot hippokampuksessa. Neuron 33, 325-340. doi: 10.1016/S0896-6273(02)00586-X
CrossRef Full Text
Buzsaki, G., and Wang, X. J. (2012). Gammaoskillaatioiden mekanismit. Annu. Rev. Neurosci. 35, 203-225. doi: 10.1146/annurev-neuro-062111-150444
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Cea-Del Rio, C. A., Lawrence, J. J., Erdelyi, F., Szabo, G. ja McBain, C. J. (2011). Kolinerginen modulaatio vahvistaa CA1-hippokampuksen kolekystokiniini-positiivisten interneuronien sisäisiä oskillaatio-ominaisuuksia. J. Physiol. 589, 609-627. doi: 10.1113/jphysiol.2010.199422
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Cea-Del Rio, C. A., Lawrence, J. J., Tricoire, L., Erdelyi, F., Szabo, G., ja McBain, C. J. (2010). M3-muskariinisen asetyylikoliinireseptorin ilmentyminen antaa erilaista kolinergistä modulaatiota neurokemiallisesti erilaisille hippokampuksen korisolujen alatyypeille. J. Neurosci. 30, 6011-6024. doi: 10.1523/JNEUROSCI.5040-09.2010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Cobb, S. R., ja Davies, C. H. (2005). Hippokampuksen solujen ja piirien kolinerginen modulointi. J. Physiol. 562, 81-88. doi: 10.1113/jphysiol.2004.076539
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Cole, A. E., ja Nicoll, R. A. (1983). Asetyylikoliini välittää hidasta synaptista potentiaalia hippokampuksen pyramidisoluissa. Science 221, 1299-1301. doi: 10.1126/science.6612345
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Davis, J. A., and Gould, T. J. (2006). DHBE: n ja MLA: n vaikutukset nikotiinin aiheuttamaan kontekstuaalisen pelon ehdollistamisen tehostumiseen C57BL/6-hiirissä. Psykofarmakologia (Berl). 184, 345-352. doi: 10.1007/s00213-005-0047-y
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Davis, J. A., ja Gould, T. J. (2009). Hippokampuksen nAChR:t välittävät nikotiinin vieroitukseen liittyviä oppimisvajauksia. Eur. Neuropsychopharmacol. 19, 551-561. doi: 10.1016/j.euroneuro.2009.02.003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Davis, J. A., Kenney, J. W., and Gould, T. J. (2007). Hippokampuksen alfa4beta2-nikotiinisen asetyylikoliinireseptorin osallistuminen akuutin nikotiinin tehostavaan vaikutukseen kontekstuaaliseen pelon ehdollistamiseen. J. Neurosci. 27, 10870-10877. doi: 10.1523/JNEUROSCI.3242-07.2007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Deiana, S., Platt, B., ja Riedel, G. (2011). Kolinerginen järjestelmä ja avaruudellinen oppiminen. Behav. Brain Res. 221, 389-411. doi: 10.1016/j.bbr.2010.11.036
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Dutar, P., Bassant, M. H., Senut, M. C. ja Lamour, Y. (1995). Septohippokampusreitti: keskeisen kolinergisen järjestelmän rakenne ja toiminta. Physiol. Rev. 75, 393-427.
Pubmed Abstract | Pubmed Full Text
Easton, A., Douchamps, V., Eacott, M., and Lever, C. (2012). Septohippokampuksen asetyylikoliinin erityinen rooli muistissa? Neuropsychologia 50, 3156-3168. doi: 10.1016/j.neuropsychologia.2012.07.022
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Fernandez De Sevilla, D., and Buno, W. (2010). NMDA- ja AMPA-reseptorivälitteisen lähetyksen muskariininen pitkäaikainen tehostaminen Schafferin kollateraalisynapseissa kehittyy erilaisten solunsisäisten mekanismien kautta. J. Neurosci. 30, 11032-11042. doi: 10.1523/JNEUROSCI.1848-10.2010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Frazier, C. J., Buhler, A. V., Weiner, J. L. ja Dunwiddie, T. V. (1998a). Alfa-bungarotoksiiniherkkien nikotiinisten asetyylikoliinireseptorien välittämät synaptiset potentiaalit rotan hippokampuksen interneuroneissa. J. Neurosci. 18, 8228-8235.
Pubmed Abstract | Pubmed Full Text
Frazier, C. J., Rollins, Y. D., Breese, C. R., Leonard, S., Freedman, R. ja Dunwiddie, T. V. (1998b). Asetyylikoliini aktivoi alfa-bungarotoksiini-herkän nikotiinivirran rotan hippokampuksen interneuroneissa, mutta ei pyramidisoluissa. J. Neurosci. 18, 1187-1195.
Pubmed Abstract | Pubmed Full Text
Freedman, R., Adler, L. E., Bickford, P., Byerley, W., Coon, H., Cullum, C. M., et al. (1994). Skitsofrenia ja nikotiinireseptorit. Harv. Rev. Psychiatry 2, 179-192. doi: 10.3109/10673229409017136
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Freund, T. F., and Buzsaki, G. (1996). Hippokampuksen interneuronit. Hippocampus 6, 347-470.
Pubmed Abstract | Pubmed Full Text
Fukudome, Y., Ohno-Shosaku, T., Matsui, M., Omori, Y., Fukaya, M., Tsubokawa, H., et al. (2004). Kaksi erilaista luokkaa muskariinien vaikutuksesta hippokampuksen estäviin synapseihin: M2-välitteinen suora tukahduttaminen ja M1/M3-välitteinen epäsuora tukahduttaminen endokannabinoidisignaalin välityksellä. Eur. J. Neurosci. 19, 2682-2692. doi: 10.1111/j.0953-816X.2004.03384.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Gahring, L. C., Persiyanov, K. ja Rogers, S. W. (2005). Hiirikantakohtaiset muutokset nikotiinireseptorin ilmentymisessä iän myötä. Neurobiol. Aging 26, 973-980. doi: 10.1016/j.neurobiolaging.2004.07.005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Gais, S., and Born, J. (2004). Matala asetyylikoliini hitaan aallon unen aikana on kriittinen deklaratiivisen muistin konsolidoinnin kannalta. Proc. Natl. Acad. Sci. U.S.A. 101, 2140-2144. doi: 10.1073/pnas.0305404101
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Giessel, A. J., ja Sabatini, B. L. (2010). M1-muskariinireseptorit lisäävät synaptisia potentiaaleja ja kalsiumin sisäänvirtausta dendriittisissä selkärangoissa estämällä postsynaptisia SK-kanavia. Neuron 68, 936-947. doi: 10.1016/j.neuron.2010.09.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Giocomo, L. M., ja Hasselmo, M. E. (2007). Glutamaatin ja asetyylikoliinin neuromodulaatio voi muuttaa piirin dynamiikkaa säätelemällä afferentin syötteen ja eksitatorisen palautteen suhteellista vaikutusta. Mol. Neurobiol. 36, 184-200. doi: 10.1007/s12035-007-0032-z
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Griguoli, M., Scuri, R., Ragozzino, D. ja Cherubini, E. (2009). Nikotiinisten asetyylikoliinireseptorien aktivointi lisää hidasta kalsiumriippuvaista kaliumkonduktanssia ja vähentää stratum oriens interneuronien ampumista. Eur. J. Neurosci. 30, 1011-1022. doi: 10.1111/j.1460-9568.2009.06914.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hasselmo, M. E. (2006). Asetyylikoliinin rooli oppimisessa ja muistissa. Curr. Opin. Neurobiol. 16, 710-715. doi: 10.1016/j.conb.2006.09.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hasselmo, M. E., and Giocomo, L. M. (2006). Kortikaalisen toiminnan kolinerginen modulointi. J. Mol. Neurosci. 30, 133-135. doi: 10.1385/JMN:30:1:133
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hasselmo, M. E., ja McGaughy, J. (2004). Korkeat asetyylikoliinitasot asettavat piirin dynamiikan huomiolle ja koodaukselle ja matalat asetyylikoliinitasot asettavat dynamiikan konsolidoinnille. Prog. Brain Res. 145, 207-231. doi: 10.1016/S0079-6123(03)45015-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hasselmo, M. E., ja Sarter, M. (2011). Kognition etuaivojen kolinergisen neuromodulaation moodit ja mallit. Neuropsychopharmacology 36, 52-73. doi: 10.1038/npp.2010.104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Jia, Y., Yamazaki, Y., Nakauchi, S., and Sumikawa, K. (2009). Alfa2-nikotiinireseptorit toimivat molekulaarisena kytkimenä, joka herättää jatkuvasti interneuronien osajoukon rotan hippokampuksen piireissä. Eur. J. Neurosci. 29, 1588-1603. doi: 10.1111/j.1460-9568.2009.06706.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Jones, S., ja Yakel, J. L. (1997). Toimivat nikotiiniset ACh-reseptorit rotan hippokampuksen interneuroneissa. J. Physiol. 504(Pt 3), 603-610. doi: 10.1111/j.1469-7793.1997.603bd.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Kellar, K. J., Whitehouse, P. J., Martino-Barrows, A. M., Marcus, K., ja Price, D. L. (1987). Muskariiniset ja nikotiiniset kolinergiset sitoutumiskohdat Alzheimerin taudin aivokuoressa. Brain Res. 436, 62-68. doi: 10.1016/0006-8993(87)91556-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Kenney, J. W., and Gould, T. J. (2008). Hippokampuksesta riippuvan oppimisen ja synaptisen plastisuuden modulointi nikotiinilla. Mol. Neurobiol. 38, 101-121. doi: 10.1007/s12035-008-8037-9
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Klausberger, T., ja Somogyi, P. (2008). Neuronaalinen monimuotoisuus ja ajallinen dynamiikka: hippokampuksen piiritoimintojen yhtenäisyys. Science 321, 53-57. doi: 10.1126/science.1149381
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Larkum, M. E., Zhu, J. J., ja Sakmann, B. (1999). Uusi solumekanismi eri aivokuoren kerroksiin saapuvien syötteiden kytkemiseksi. Nature 398, 338-341. doi: 10.1038/18686
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Lawrence, J. J., Statland, J. M., Grinspan, Z. M. ja McBain, C. J. (2006). Muskariinisen signaloinnin solutyyppikohtainen riippuvuus hiiren hippokampuksen stratum oriens interneuroneissa. J. Physiol. 570, 595-610. doi: 10.1113/jphysiol.2005.100875
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Leonard, S., Adams, C., Breese, C. R., Breese, C. R., Adler, L. E., Bickford, P., Byerley, W., ym. et al. (1996). Nikotiinireseptorien toiminta skitsofreniassa. Schizophr. Bull. 22, 431-445. doi: 10.1093/schbul/22.3.431
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Levin, E. D. (2002). Nikotiinireseptorin alatyypit ja kognitiivinen toiminta. J. Neurobiol. 53, 633-640. doi: 10.1002/neu.10151
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Levin, E. D., Bradley, A., Addy, N. ja Sigurani, N. (2002). Hippokampuksen alfa 7 ja alfa 4 beeta 2 nikotiinireseptorit ja työmuisti. Neuroscience 109, 757-765. doi: 10.1016/S0306-4522(01)00538-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Levin, E. D., Petro, A., Rezvani, A. H., Pollard, N., Christopher, N. C., Strauss, M., ym. et al. (2009). Nikotiinisen alfa7- tai beeta2-reseptorin tyrmäys: vaikutukset radiaalisen käden labyrintin oppimiseen ja pitkäaikaiseen nikotiinin kulutukseen hiirissä. Behav. Brain Res. 196, 207-213. doi: 10.1016/j.bbr.2008.08.048
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
McQuiston, A. R., ja Madison, D. V. (1999a). Muskariinireseptorin aktiivisuudella on useita vaikutuksia CA1-hippokampuksen interneuronien lepokalvopotentiaaliin. J. Neurosci. 19, 5693-5702.
Pubmed Abstract | Pubmed Full Text
McQuiston, A. R., ja Madison, D. V. (1999b). Muskariinireseptorin aktiivisuus indusoi jälkidepolarisaation hippokampuksen CA1 interneuronien osapopulaatiossa. J. Neurosci. 19, 5703-5710.
Pubmed Abstract | Pubmed Full Text
McQuiston, A. R., ja Madison, D. V. (1999c). Nikotiinireseptorin aktivaatio herättää erilaisia interneuronien alatyyppejä rotan hippokampuksessa. J. Neurosci. 19, 2887-2896.
Pubmed Abstract | Pubmed Full Text
Miles, R., Toth, K., Gulyas, A. I., Hajos, N. ja Freund, T. F. (1996). Erot somaattisen ja dendriittisen eston välillä hippokampuksessa. Neuron 16, 815-823. doi: 10.1016/S0896-6273(00)80101-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Nagode, D. A., Tang, A. H., Karson, M. A., Klugmann, M. ja Alger, B. E. (2011). ACh: n optogeneettinen vapautuminen indusoi perisomaattisten IPSC: iden rytmisiä purskeita hippokampuksessa. PLoS ONE 6:e27691. doi: 10.1371/journal.pone.0027691
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Nagode, D. A., Tang, A. H., Yang, K., and Alger, B. E. (2014). Optogeneettinen identifiointi sisäisen kolinergisesti ohjatun inhiboivan oskillaattorin tunnistaminen, joka on herkkä kannabinoideille ja opioideille hippokampuksen CA1: ssä. J. Physiol. 592, 103-123. doi: 10.1113/jphysiol.2013.257428
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Nott, A., ja Levin, E. D. (2006). Dorsaalisen hippokampuksen alfa7- ja alfa4beta2-nikotiinireseptorit ja muisti. Brain Res. 1081, 72-78. doi: 10.1016/j.brainres.2006.01.052
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Parra, P., Gulyas, A. I., and Miles, R. (1998). Kuinka monta estävien solujen alatyyppiä hippokampuksessa? Neuron 20, 983-993. doi: 10.1016/S0896-6273(00)80479-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Perry, D. C., Davila-Garcia, M. I., Stockmeier, C. A., and Kellar, K. J. (1999). Lisääntyneet nikotiinireseptorit tupakoitsijoiden aivoissa: kalvon sitoutumis- ja autoradiografiatutkimukset. J. Pharmacol. Exp. Ther. 289, 1545-1552.
Pubmed Abstract | Pubmed Full Text
Pitler, T. A., and Alger, B. E. (1992). GABAergisten interneuronien kolinerginen heräte rotan hippokampusviipaleessa. J. Physiol. 450, 127-142.
Pubmed Abstract | Pubmed Full Text
Power, A. E., Vazdarjanova, A., ja McGaugh, J. L. (2003). Muskariiniset kolinergiset vaikutukset muistin konsolidoinnissa. Neurobiol. Learn. Mem. 80, 178-193. doi: 10.1016/S1074-7427(03)00086-8
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Simon, A. P., Poindessous-Jazat, F., Dutar, P., Epelbaum, J. ja Bassant, M. H. (2006). Anatomisesti tunnistettujen neuronien laukaisuominaisuudet nukutettujen ja nukuttamattomien hillittyjen rottien mediaalisessa septumissa. J. Neurosci. 26, 9038-9046. doi: 10.1523/JNEUROSCI.1401-06.2006
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Smiley, J. F., Morrell, F., ja Mesulam, M. M. (1997). Kolinergiset synapsit ihmisen aivokuoressa: ultrastruktuuritutkimus sarjapoikkileikkauksissa. Exp. Neurol. 144, 361-368. doi: 10.1006/exnr.1997.6413
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Stone, T. W. (2007). Kynureniinihappo estää nikotiinisen synaptisen siirron hippokampuksen interneuroneihin nuorilla rotilla. Eur. J. Neurosci. 25, 2656-2665. doi: 10.1111/j.1460-9568.2007.05540.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Storm-Mathisen, J. (1970). Asetyylikoliiniesteraasin kvantitatiivinen histokemia rotan hippokampuksen alueella korreloituna histokemialliseen värjäykseen. J. Neurochem. 17, 739-750. doi: 10.1111/j.1471-4159.1970.tb03344.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Sudweeks, S. N., ja Yakel, J. L. (2000). Neuronaalisten nikotiinisten ACh-reseptorien toiminnallinen ja molekulaarinen karakterisointi rotan CA1-hippokampuksen neuroneissa. J. Physiol. 527(Pt 3), 515-528. doi: 10.1111/j.1469-7793.2000.00515.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Szabo, G. G., Holderith, N., Gulyas, A. I., Freund, T. F., and Hajos, N. (2010). Perisomaattisten inhibitoristen solutyyppien erilaiset synaptiset ominaisuudet ja niiden erilainen modulointi kolinergisen reseptorin aktivoinnilla hiiren hippokampuksen CA3-alueella. Eur. J. Neurosci. 31, 2234-2246. doi: 10.1111/j.1460-9568.2010.07292.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Tang, A. H., Karson, M. A., Nagode, D. A., McIntosh, J. M., Uebele, V. N., Renger, J. J. ym. et al. (2011). Hermopäätteen nikotiiniset asetyylikoliinireseptorit käynnistävät kvantaalisen GABA:n vapautumisen perisomaattisista interneuroneista aktivoimalla aksonaalisia T-tyypin (Cav3) Ca(2)(+)-kanavia ja Ca(2)(+)-vapautumista varastoista. J. Neurosci. 31, 13546-13561. doi: 10.1523/JNEUROSCI.2781-11.2011
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Teles-Grilo Ruivo, L. M., ja Mellor, J. R. (2013). Hippokampusverkon toiminnan kolinerginen modulointi. Front. Synaptic Neurosci. 5:2. doi: 10.3389/fnsyn.2013.00002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Tsubokawa, H. (2000). Na+-piikin takaisinkulkeutumisen hallinta solunsisäisellä signaloinnilla pyramidineuronin dendriiteissä. Mol. Neurobiol. 22, 129-141. doi: 10.1385/MN:22:1-3:129
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Tsubokawa, H., ja Ross, W. N. (1997). Muskariininen modulaatio piikin takaisinkulkeutumisesta hippokampuksen CA1-pyramidineuronien apikaalisissa dendriiteissä. J. Neurosci. 17, 5782-5791.
Pubmed Abstract | Pubmed Full Text
Turrini, P., Casu, M. A., Wong, T. P., De, K. Y., Ribeiro-Da-Silva, A. ja Cuello, A. C. (2001). Kolinergiset hermopäätteet muodostavat klassisia synapseja rotan aivokuorella: synaptinen malli ja ikään liittyvä atrofia. Neuroscience 105, 277-285. doi: 10.1016/S0306-4522(01)00172-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Umbriaco, D., Garcia, S., Beaulieu, C. ja Descarries, L. (1995). Asetyylikoliinin, noradrenaliinin, serotoniinin ja GABA:n aksonipäätteiden suhteelliset ominaisuudet aikuisen rotan hippokampuksen stratum radiatumissa (CA1). Hippocampus 5, 605-620. doi: 10.1002/hipo.450050611
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Umbriaco, D., Watkins, K. C., Descarries, L., Cozzari, C. ja Hartman, B. K. (1994). Aikuisen rotan parietaalisen aivokuoren asetyylikoliini-innervaation ultrastruktuuriset ja morfometriset piirteet: elektronimikroskooppinen tutkimus sarjapoikkileikkauksissa. J. Comp. Neurol. 348, 351-373. doi: 10.1002/cne.903480304
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Vizi, E. S., Fekete, A., Karoly, R. ja Mike, A. (2010). Aivotoimintoihin osallistuvat ei-synaptiset reseptorit ja kuljettajat ja lääkehoidon kohteet. Br. J. Pharmacol. 160, 785-809. doi: 10.1111/j.1476-5381.2009.00624.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Widmer, H., Ferrigan, L., Davies, C. H., ja Cobb, S. R. (2006). Evokoidut hitaat muskariiniset asetyylikoliinergiset synaptiset potentiaalit rotan hippokampuksen interneuroneissa. Hippocampus 16, 617-628. doi: 10.1002/hipo.20191
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Wu, J., Kuo, Y. P., George, A. A.., Xu, L., Hu, J. ja Lukas, R. J. (2004). beeta-amyloidi estää suoraan ihmisen alfa4beta2-nikotiinisia asetyylikoliinireseptoreita, jotka on heterologisesti ilmaistu ihmisen SH-EP1-soluissa. J. Biol. Chem. 279, 37842-37851. doi: 10.1074/jbc.M400335200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Zhang, H., Lin, S. C., and Nicolelis, M. A. (2010). Hippokampuksen asetyylikoliinin vapautumisen ja theta-oskillaatioiden välinen spatiotemporaalinen kytkentä in vivo. J. Neurosci. 30, 13431-13440. doi: 10.1523/JNEUROSCI.1144-10.2010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text