FunctionEdit
AzABA eredetileg úgy gondolták, hogy részt vesz az abszcisszióban, innen kapta a nevét. Ma már tudjuk, hogy ez csak kevés növény esetében van így. Az ABA által közvetített jelátvitel a környezeti stresszre és a növényi kórokozókra adott növényi válaszokban is fontos szerepet játszik. Az ABA bioszintézisének növényi génjeit és az útvonal szekvenciáját már feltárták. Az ABA-t egyes növénypatogén gombák is termelik, a növényi ABA bioszintézisétől eltérő bioszintetikus úton.
A télre való felkészülés során az ABA a végrügyekben termelődik. Ez lelassítja a növény növekedését, és a levélprimordiumokat a pikkelyek fejlődésére irányítja, hogy a hideg évszak alatt megvédjék a nyugvó rügyeket. Az ABA gátolja a sejtek osztódását is az érrendszeri kambiumban, alkalmazkodva a téli hideg körülményekhez az elsődleges és másodlagos növekedés felfüggesztésével.
Az abszcizinsav a gyökerekben is termelődik a talaj vízpotenciáljának csökkenésére (ami a száraz talajjal jár együtt) és más olyan helyzetekre válaszul, amelyekben a növény stressznek lehet kitéve. Az ABA ezután transzlokálódik a levelekbe, ahol gyorsan megváltoztatja a sztómák védősejtjeinek ozmotikus potenciálját, aminek következtében azok összezsugorodnak és a sztómák bezáródnak. Az ABA által kiváltott sztómazárás csökkenti a transzspirációt (a víz párolgását a sztómákból), így megakadályozza a további vízveszteséget a levelekből az alacsony vízellátottság idején. Szoros lineáris korrelációt találtak a levelek ABA-tartalma és a levelek vezetőképessége (sztómás ellenállás) között a levélfelület alapján.
A magok csírázását gátolja az ABA a gibberellinnel antagonizmusban. Az ABA megakadályozza a magok dormánságának elvesztését is.
A Nottinghami Arabidopsis Állományközpontban számos ABA-mutáns Arabidopsis thaliana növényt azonosítottak és állnak rendelkezésre – mind az ABA-termelésben hiányos, mind a hatásával szemben megváltozott érzékenységű növényeket. Az ABA-ra túlérzékeny vagy érzéketlen növények fenotípusokat mutatnak a magok nyugalmi állapotában, a csírázásban, a sztómák szabályozásában, és egyes mutánsok csonka növekedést és barna/sárga leveleket mutatnak. Ezek a mutánsok tükrözik az ABA fontosságát a magcsírázásban és a korai embriófejlődésben.
A pirabaktin (egy piridil tartalmú ABA aktivátor) egy naftalin-szulfonamid hipokotil sejtterjedést gátló anyag, amely a mag ABA jelátviteli útvonalának agonistája. Ez az ABA útvonal első olyan agonistája, amely szerkezetileg nem kapcsolódik az ABA-hoz.
HomeosztázisSzerkesztés
BioszintézisSzerkesztés
Az abszcizinsav (ABA) egy izoprenoid növényi hormon, amely a plasztidális 2-C-metil-D-eritritol-4-foszfát (MEP) útvonalon szintetizálódik; a szerkezetileg rokon szeszkviterpénekkel ellentétben, amelyek a mevalonsavból származó farnezil-difoszfát (FDP) prekurzorból keletkeznek, az ABA C15 gerincét a MEP-ben lévő C40 karotinoidok hasadása után képzik. A zeaxantin az első elkötelezett ABA prekurzor; enzimkatalizált epoxidációk és izomerizációk sorozata a violaxantinon keresztül, és a C40 karotinoid végső hasadása egy diooxigenizációs reakcióval adja a proximális ABA prekurzort, a xantoxint, amely aztán tovább oxidálódik ABA.-vá az abszcizinaldehidon keresztül.

Abamint tervezték, szintetizálták, fejlesztették, majd szabadalmaztatták, mint az első specifikus ABA bioszintézis gátlót, amely lehetővé teszi az ABA endogén szintjének szabályozását.
Az ABA bioszintézis helye és időzítéseSzerkesztés
- A vegetatív szövetek kiszáradásakor és a gyökerek talajtömörödésekor szabadul fel.
- Szintetizálódik a zöld termésekben a téli időszak kezdetén
- Szintetizálódik az érő magvakban, létrehozva a nyugalmi állapotot
- Mobil a levélben és gyorsan transzlokálódhat a gyökerekből a levelekbe a xilém transzspirációs áramlásával
- Elterjed a környezeti stresszre, pl. hő-, víz-, sóstresszre válaszul
- Szintetizálódik minden növényi részben, pl, gyökerek, virágok, levelek és szárak
- ABA szinte minden olyan sejtben szintetizálódik, amely kloroplasztiszokat vagy amiloplasztiszokat tartalmaz
InaktiválásSzerkesztés
ABA a CYP707A (a P450 enzimek egy csoportja) segítségével fáziasavvá katabolizálható vagy az AOG enzimen keresztül glükózkonjugációval (ABA-glükózészter) inaktiválható. A CYP707A-kon keresztül történő katabolizmus nagyon fontos az ABA homeosztázis szempontjából, és e gének mutánsai általában magasabb ABA-szintet akkumulálnak, mint az ABA bioszintetikus géneket túlreprezentáló vonalak. Talajbaktériumokban egy alternatív katabolikus útvonalról számoltak be, amely a vomifoliol-dehidrogenáz enzimen keresztül dehidrovomifoliolhoz vezet.
HatásokSzerkesztés
- Antitranszpiráns – Indukálja a sztómák záródását, csökkentve a transzspirációt a vízveszteség megelőzése érdekében.
- Gátolja a gyümölcsök érését
- A sejtnövekedés gátlásával felelős a magok nyugalmi állapotáért – gátolja a magok csírázását
- Gátolja a kinetin nukleotid szintézisét
- Leregulálja a fotoszintézishez szükséges enzimeket.
- Az endodermiszre hatva megakadályozza a gyökerek növekedését, ha sós körülményeknek van kitéve
- Késlelteti a sejtosztódást
- Dormancia indukáló – A vetőmagok nyugalmi állapotának előidézésére használják.
- Transzpirációgátlóként használják – A szárazságra hajlamos területeken a vízstressz komoly probléma a mezőgazdasági termelésben. ezért javasolják az ABA permetezését, amely néhány napra a sztómák részleges záródását okozza , hogy csökkentse a transzpirációs vízveszteséget
SzignálkaszkádSzerkesztés
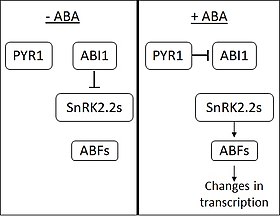
ABA hiányában, az ABI1-INSENSITIVE1 (ABI1) foszfatáz gátolja az SNF1-hez kapcsolódó fehérje kinázok (2. alcsalád) (SnRK2s) működését. Az ABA-t a PYRABACTIN RESISTANCE 1 (PYR1) és a PYR1-szerű membránfehérjék érzékelik. Az ABA megkötésekor a PYR1 az ABI1-hez kötődik és gátolja azt. Amikor az SnRK2 felszabadul a gátlás alól, az ABA RESPONSIVE ELEMENT-BINDING FACTOR (ABF) család számos transzkripciós faktorát aktiválja. Az ABF-ek ezután számos gén kifejeződésében okoznak változásokat. Úgy gondolják, hogy a növényi gének mintegy 10%-át az ABA szabályozza.