A mechanizmus jobb megértéséhez az alábbiakban a szaglás útvonalának egyszerű lebontása következik. Amikor az ember rág, az illékony ízvegyületek átjutnak a nasopharanxon és a szaglóreceptorokon.
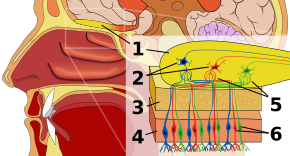
SzaglóhámSzerkesztés
A szaglórendszer első állomása a szaglóhám, vagyis az orrüreg tetején nyugvó szövet, amely a szaglóreceptoroknak ad otthont. A szagreceptorok bipoláris neuronok, amelyek szaganyagokat kötnek meg a levegőből, és a szaglóidegben gyűlnek össze, mielőtt axonjaik a szaglógumóban lévő mitrális sejtek dendritjeihez jutnak. A szájban és az orrban lévő érzékelő receptorok nyugalmi állapotban polarizáltak, és a környezet valamilyen változására, például a szagmolekulákkal való érintkezésre reagálva depolarizálódnak. A funkcionális csoportokkal rendelkező szénhidrogénláncokból álló szagmolekulák az orrban és a szájban lévő érzékszervi receptorokhoz kötődnek. A funkcionális csoportok tulajdonságai közé tartoznak: (1) a szénlánc hossza, (2) a végcsoport, amely egybeesik a különböző szagokhoz kapcsolódó különbségekkel, (3) az oldalcsoport, (4) a kiralitás, (5) az alak és (6) a méret. Amikor a szagmolekulák az érzékszervi receptorokhoz kötődnek, azt e tulajdonságok szerint teszik. Minden szaglósejtnek egyetlen receptortípusa van, de ez a receptor “széleskörűen hangolható”, és a szagmolekulák további kölcsönhatásba lépnek a receptorok szintjén, ami azt jelenti, hogy bizonyos esetekben előfordulhat, hogy egy szagmolekula önmagában nem kötődik egy receptorhoz, de egy másik szagmolekula jelenlétében az eredeti kötődne, és így csak a második molekula jelenlétében okozna szagérzetet.
SzaglógumóSzerkesztés
A szaglógumóban a szagmolekulák térbeli leképezése történik. Ezeket a térbeli ábrázolásokat “szagképeknek” nevezzük. A térbeli reprezentáció lehetővé teszi a laterális gátlást, illetve a kontrasztfokozást és az erősítés tömörítését. A kontrasztfokozás érzékeny a változásra, és kiemeli azokat az ingereket az agyban, amelyek inkább változnak, mint nyugalomban vannak. Az erősítéskompresszió fokozza az alacsony intenzitású ingerekre való érzékenységet, miközben csökkenti a magas intenzitású ingerekre való érzékenységet. A szaglógumó, bár a kutatók még mindig a megértés elsődleges szakaszában vannak, megkülönbözteti a szaglást a többi érzékszervtől, mert eltérést jelent az érzékelési útvonalban attól, ami az összes többi érzékszervre jellemző. Ugyanis minden nem szaglással kapcsolatos érzékszervi információ a receptorszint után a talamuszon halad át, de az a tény, hogy a szaginformáció ehelyett egy saját, erre specializált területre kerül, a szaglás primitív történetére és/vagy a szaginformációnak az agykéreg felé vezető úton történő külön típusú feldolgozására utalhat. A szaglógumóban glomerulusok, vagyis sejtcsomópontok találhatók, amelyeken a mitrális sejteken kívül több ezer azonos típusú receptor fut össze. Ez a szerveződés lehetővé teszi, hogy hatalmas mennyiségű információ tömören reprezentálható legyen anélkül, hogy ugyanolyan nagyszámú receptortípusra lenne szükség. A szaginformációk eredő kombinációját a szaglógumó szintjén szagképnek nevezik.
Képalkotás a szaglógumóbanSzerkesztés
2DG módszerSzerkesztés
1977-ben Lou Sokoloff, Seymour Kety és Floyd E. Bloom biokémikusok kifejlesztettek egy módszert az agyi aktivitás feltérképezésére azáltal, hogy nyomon követték a patkányagy oxigén metabolizmusát. Az idegsejteknek oxigénre és glükózra van szükségük az energiához. A 2-deoxiglükóz (2DG) egy radioaktív glükózizotóp, amelyet nyomon lehet követni az agyban, mivel nyomot hagy a sejtben, ahol normális esetben glükózként energiává metabolizálódna. A sejtek egy bizonyos régiójának stimulálása után röntgenfelvételeket lehet szeletelni, amelyekből kiderül, hogy mely sejtek voltak aktívak, különösen a szinaptikus csomópontoknál.
A funkcionális mágneses rezonancia képalkotás (fMRI) szintén használható a szaganyagcsere mérésére. Ez a módszer nem terminális, mint a 2-deoxiglükóz módszer, így egy állatot több szaggal is meg lehet mérni, és az így kapott képeket össze lehet hasonlítani.
Zöld fluoreszcens fehérje módszerSzerkesztés
A zöld fluoreszcens fehérje módszerrel végül az egereket genetikailag úgy alakítják át, hogy egy fehérjét expresszáljanak az aktív neuronokban, majd az egér koponyájában elhelyezett kamerával mérhető az aktivitás.
EredményekSzerkesztés
Ezek a módszerek legfőképpen azt mutatják, hogy a szaglási információ szerveződése a szaglógumóban térbeli. Hasonló molekuláris mintázatok hasonló aktivációs mintázatokat eredményeznek a glomerulusok tekintetében, és az egymáshoz közelebb elhelyezkedő glomerulusok a szaginformáció hasonló jellemzőit kódolják.
SzaglókéregSzerkesztés
A háromrétegű, piramissejteket tartalmazó szaglókéreg a következő viszonyítási pont a szaglási útvonalon. Egy-egy piramissejt a szaglógumóból származó mitrális sejtek sokaságától kapja az információt, így a szaglókéregben eloszlik a korábban szervezett glomeruláris mintázat. A mitrális sejtek információinak ez a szétszóródása lehetővé teszi az öngerjesztő visszacsatolási kapcsolatokat, az oldalirányú gerjesztést, valamint az ön- és oldalirányú gátlást. Ezek a folyamatok hozzájárulnak a Hebb-tanuláshoz, amelyet Donald O. Hebbről neveztek el, és gyakran leegyszerűsítve a “neuronok, amelyek együtt tüzelnek, együtt drótoznak” mondással jellemzik. A hosszú távú potenciálás, a Hebb-tanulás idegi mechanizmusa lehetővé teszi a memóriaképzést a piramissejtek szintjén. A Hebb-tanulás tehát lényegében az a jelenség, amelynek révén a szaglóagy “megjegyzi” a szagmolekulák kombinációinak kimenetét, és lehetővé teszi a korábban érzékelt kombinációk gyorsabb felismerését, mint az újakét azáltal, hogy a tárolt bemenethez igazítja őket. Az így kapott szagokat, amelyeket korábban szagképeknek neveztek, a felismerés céljából a szaglóagykéreg tárolja, mostantól szagobjektumoknak nevezzük. A tapasztalat tehát erősíti a jel-zaj arányt, amennyiben egy korábban érzékelt szagobjektum könnyebben megkülönböztethető a nagyobb háttérzajjal szemben.
Orbitofrontális kéregSzerkesztés
Az orbitofrontális kéreg (OFC) a szaginformációk végső célpontja, és itt keletkezik a tudatos szagérzékelés. A szaginformáció közvetlenül a szaglóérző kéregen való áthaladás után jut be, ami megkülönbözteti a többi érzékszervi információtól, amelyek először a talamuszon haladnak át. Az OFC a prefrontális kéreg dorzális részén helyezkedik el, így a szaginformáció közvetlenül a prefrontális kéregbe, vagyis az agy fő döntéshozó területére jut. Mindössze háromféle neuronon haladnak át a szagmolekulák, mielőtt az OFC-be érkeznek: a szaglóhámban lévő szagló receptorsejteken, a mitrális sejteken és a szagló piramis neuronokon.
Az OFC szintjén más agyterületekkel is történnek asszociációk, beleértve a szájból érkező inputot (szomatoszenzáció), az érzelmi inputot (amygdala), a vizuális információt és az értékelő információt (prefrontális kéreg). Az OFC felelős a szelektív szaghangolásért, az érzékszervi területek összeolvadásáért és a szagok hedonikus értékeléséért.