- Introduzione
- MS/DBB Attività dei neuroni colinergici e rilascio di acetilcolina nell’ippocampo CA1
- Effetti dell’attivazione dei recettori muscarinici sugli interneuroni inibitori dell’ippocampo CA1
- Attivazione dei recettori muscarinici negli interneuroni ippocampali CA1 in seguito al rilascio di acetilcolina
- Effetti dell’attivazione dei recettori nicotinici sugli interneuroni ippocampali CA1
- Attivazione dei recettori nicotinici negli interneuroni ippocampali CA1 in seguito al rilascio di acetilcolina
- Effetti del rilascio di acetilcolina sulla funzione della rete ippocampale CA1 dal punto di vista del potenziale di membrana degli interneuroni
- Conflict of Interest Statement
- Riconoscimento
Introduzione
L’acetilcolina viene rilasciata in tutto il sistema nervoso centrale (SNC) dei mammiferi, dove ha un impatto sulle funzioni cerebrali globali influenzando i cicli sonno-veglia, l’attenzione e la formazione della memoria. Una regione del cervello fortemente innervata da afferenze colinergiche dal setto mediale e dalla banda diagonale del complesso di Broca (MS/DBB) è l’ippocampo (Dutar et al., 1995). Funzionalmente, il rilascio di acetilcolina nell’ippocampo è stato proposto per aiutare la formazione o il recupero dei ricordi a seconda della concentrazione extracellulare di acetilcolina (Power et al., 2003; Hasselmo e Giocomo, 2006; Kenney e Gould, 2008; Deiana et al., 2011; Hasselmo e Sarter, 2011; Easton et al., 2012; Blake et al., 2014). Il meccanismo con cui i terminali colinergici MS/DBB influenzano la funzione della rete ippocampale è attraverso l’attivazione dei recettori muscarinici e nicotinici situati sui dendriti, corpi cellulari e terminali assonali dei neuroni piramidali e interneuroni inibitori, così come sugli astrociti (Cobb e Davies, 2005; Teles-Grilo Ruivo e Mellor, 2013). Anche se l’acetilcolina agisce su più siti e su diversi tipi di cellule, una parte della sua influenza deriva probabilmente dai suoi effetti sulla funzione degli interneuroni.
Gli interneuroni inibitori svolgono un ruolo cruciale nell’elaborazione delle informazioni nell’ippocampo. Gli interneuroni sono molto diversi nella struttura anatomica e nella funzione presunta (Freund e Buzsaki, 1996; Klausberger e Somogyi, 2008). A seconda del sottotipo di interneurone e dove innerva la cellula piramidale, un singolo interneurone può bloccare completamente l’attività in un dendrite, cambiare la fase di sparo del potenziale d’azione al soma, o impedire completamente lo sparo del potenziale d’azione al corpo della cellula piramidale (Miles et al., 1996; Larkum et al., 1999). A livello di rete, gli interneuroni contribuiscono alla generazione di attività sincrona tra popolazioni di neuroni principali ad una varietà di frequenze comportamentalmente rilevanti (Buzsaki, 2002; Buzsaki e Wang, 2012). Dato l’impatto significativo che i singoli interneuroni hanno sulla funzione della rete neuronale, è probabile che una parte considerevole dell’influenza dell’acetilcolina sull’attività ippocampale derivi dalla modulazione degli interneuroni. Anche se i recettori colinergici hanno dimostrato di influenzare i terminali presinaptici inibitori (Behrends e Ten Bruggencate, 1993; Tang et al., 2011) e l’eccitabilità degli interneuroni (McQuiston e Madison, 1999b; Griguoli et al., 2009; Cea-Del Rio et al, 2010, 2011), questa minireview limiterà la sua attenzione a studi recenti che hanno indagato l’effetto del rilascio di acetilcolina sui cambiamenti nel potenziale di membrana interneurone, in particolare nella CA1 ippocampale.
MS/DBB Attività dei neuroni colinergici e rilascio di acetilcolina nell’ippocampo CA1
L’impatto che il rilascio di acetilcolina ha nell’ippocampo CA1 e la misura in cui i diversi sottotipi di interneuroni sono interessati dipenderà dalla posizione specifica e dalla densità dei terminali degli assoni colinergici e dal suo enzima inattivante, l’acetilcolinesterasi. In particolare, sia le fibre colinergiche che l’acetilcolinesterasi hanno dimostrato di essere distribuite in modo differenziato tra gli strati dell’ippocampo CA1. Nel topo, è stato dimostrato che le fibre colinergiche sono distribuite uniformemente tranne che per due bande di maggiore densità nello strato pyamidale (SP) e al confine tra lo strato radiatum (SR) e lo strato lacunoso-molecolare (SLM) (Aznavour et al., 2002). Nel ratto, simili bande a densità più elevata sono state osservate nel SP e al confine tra SR e SLM. Tuttavia, rispetto allo strato oriens (SO), sono state osservate densità inferiori nel SR e densità ancora più basse nel SLM (SO > SR > SLM) (Aznavour et al., 2002). La distribuzione dell’acetilcolinesterasi nell’ippocampo CA1 è complementare a quella dell’input colinergico, con densità più elevate osservate tra SP e SO e un altro picco nella SLM vicino al confine con la SR (Storm-Mathisen, 1970). Coerentemente con questi dati anatomici, le misure di aumento del rilascio di acetilcolina durante i ritmi theta hanno dimostrato che le concentrazioni di acetilcolina erano più alte vicino allo strato piramidale (Zhang et al., 2010). Questa distribuzione differenziale delle fibre colinergiche e dei livelli extracellulari di acetilcolina è particolarmente importante se si considera che non tutti i terminali colinergici nell’ippocampo sembrano trasmettere sinapticamente l’acetilcolina. Sia nell’ippocampo che nella neocorteccia, è stato stimato che l’85-93% dei terminali assonici colinergici non hanno specializzazione postsinaptica e quindi la maggior parte dei terminali colinergici sono stati proposti per trasmettere l’acetilcolina per volume o per trasmissione non sinaptica (Umbriaco et al., 1994, 1995). Tuttavia, altri gruppi hanno stimato che la maggior parte dei terminali colinergici (66-67%) nella neocorteccia fanno connessioni sinaptiche classiche (Smiley et al., 1997; Turrini et al., 2001). Indipendentemente da questa discrepanza, una parte significativa dei terminali sembra rilasciare acetilcolina nello spazio extracellulare in modo paracrino. Ciò richiede che l’acetilcolina rilasciata terminalmente diffonda distanze significative oltre l’acetilcolinesterasi per legarsi ai recettori sugli elementi postsinaptici. Così, regioni o strati con densità favorevoli di terminali colinergici (più alti) e/o acetilcolinesterasi (più bassi) possono risultare in maggiori concentrazioni extracellulari di acetilcolina che possono essere più efficaci nel trasmettere l’acetilcolina attraverso la trasmissione del volume. Inoltre, è possibile che ci sia un sottoinsieme di terminali che sono più attivi, hanno una maggiore probabilità di rilascio o possono rilasciare più neurotrasmettitore. Questi terminali possono essere più efficaci nel mediare la trasmissione del volume e nell’influenzare gli interneuroni inibitori vicini.
Il rilascio di acetilcolina dai terminali colinergici dipende dall’attività dei neuroni colinergici nella MS/DBB. Tuttavia, i modelli di accensione dei neuroni colinergici della SM/DBB riportati in letteratura hanno mostrato una certa variabilità (Barrenechea et al., 1995; Brazhnik e Fox, 1997, 1999; Simon et al., 2006). Un piccolo numero di neuroni colinergici MS/DBB anatomicamente identificati, registrati in roditori svegli e immobilizzati, sono stati segnalati per avere bassi tassi di cottura irregolari (<2 Hz) (Simon et al., 2006). Al contrario, neuroni anatomicamente non identificati con forme d’onda del potenziale d’azione coerente con i neuroni colinergici MS/DBB sono stati segnalati per sparare a tassi fino a 30 Hz (Brazhnik e Fox, 1999). Quindi, rimane poco chiaro quali siano le frequenze che meglio descrivono i modelli di sparo dei neuroni colinergici nella MS/DBB o se essi cadono lungo un ampio continuum. Tuttavia, potenziali differenze nella frequenza di accensione o nella durata dell’attività dei neuroni colinergici potrebbero avere effetti variabili sui diversi sottotipi di interneuroni attraverso differenze locali nelle concentrazioni di acetilcolina.
Effetti dell’attivazione dei recettori muscarinici sugli interneuroni inibitori dell’ippocampo CA1
L’interruzione della funzione colinergica del MS/DBB mediante blocco sistemico dei recettori muscarinici o iniezione diretta di antagonisti dei recettori muscarinici nell’ippocampo può compromettere la memoria e la codifica delle informazioni spaziali (Blokland et al, 1992; Atri et al., 2004; Hasselmo, 2006). Un ruolo potenziale per gli interneuroni inibitori nella modulazione dei recettori muscarinici della funzione ippocampale era inizialmente basato sulle osservazioni che l’applicazione esogena di agonisti colinergici ha portato ad un aumento delle correnti postsinaptiche inibitorie spontanee (sIPSCs) nei neuroni piramidali CA1 (Pitler e Alger, 1992). Questi dati hanno indirettamente suggerito che un sottoinsieme di interneuroni inibitori può essere depolarizzato dall’attivazione dei recettori muscarinici e sono stati successivamente confermati da registrazioni dirette (Parra et al., 1998; McQuiston e Madison, 1999a). Tuttavia, non tutti gli interneuroni hanno risposto all’attivazione del recettore muscarinico depolarizzandosi. Alcuni interneuroni erano iperpolarizzati o mostravano risposte bifasiche, e alcuni non rispondevano all’applicazione esogena dell’agonista muscarinico (Parra et al., 1998; McQuiston e Madison, 1999a). Inoltre, ogni tipo di risposta muscarinica non poteva essere correlato con un sottotipo morfologico di interneurone. Questi risultati sono stati ulteriormente complicati dall’osservazione che i recettori muscarinici possono inibire il rilascio di GABA da un sottogruppo di interneuroni inibitori perisomatici (Behrends e Ten Bruggencate, 1993; Fukudome et al., 2004; Szabo et al., 2010) e l’attivazione dei recettori muscarinici può aumentare l’eccitabilità degli interneuroni attraverso la generazione di depolarizzazioni successive (McQuiston e Madison, 1999b; Lawrence et al., 2006). Così, l’impatto che il rilascio di acetilcolina ha sulla popolazione di interneuroni è complesso e risulta nel reclutamento di alcuni interneuroni mentre ne inibisce altri.
Attivazione dei recettori muscarinici negli interneuroni ippocampali CA1 in seguito al rilascio di acetilcolina
Anche se le risposte sinaptiche colinergiche muscariniche sono state misurate per la prima volta nel 1983 nei neuroni piramidali CA1 (Cole e Nicoll, 1983), solo nel 2006 sono state misurate le risposte muscariniche al rilascio di acetilcolina evocato elettricamente negli interneuroni inibitori ippocampali CA1 (Widmer et al., 2006). Questo studio ha dimostrato che l’acetilcolina rilasciata terminalmente aveva effetti divergenti su diversi sottotipi di interneuroni. Gli interneuroni potevano rispondere depolarizzandosi, iperpolarizzandosi o con risposte bifasiche. Nel complesso, la maggior parte degli interneuroni che rispondevano producevano depolarizzazioni (64%) mentre le iperpolarizzazioni erano osservate raramente (13%) (Widmer et al., 2006). Inoltre, come studi precedenti che utilizzavano l’applicazione esogena di agonisti muscarinici (Parra et al., 1998; McQuiston e Madison, 1999a), i diversi tipi di risposta muscarinica evocati elettricamente non potevano essere correlati a specifici sottotipi anatomici di interneuroni (Widmer et al., 2006). Questi risultati sono stati recentemente confermati da studi optogenetici utilizzando il rilascio evocato in risposta alla luce-attivazione (Nagode et al., 2011; Bell et al., 2013). Tuttavia, in uno di questi studi optogenetici, interneuroni che rispondono con risposte muscariniche bifasiche (25%), iperpolarizzanti (35%), e depolarizzanti (40%) erano più equamente distribuiti tra i diversi tipi di risposta (Bell et al., 2013). È importante notare che l’acetilcolina rilasciata optogeneticamente ha prodotto prevalentemente risposte muscariniche (80%) rispetto alle risposte nicotiniche (17%). Il restante 3% degli interneuroni rispondenti aveva entrambe le risposte muscariniche e nicotiniche. Inoltre, le iperpolarizzazioni muscariniche erano mediate dall’attivazione dei recettori M4 mentre le depolarizzazioni erano probabilmente prodotte dall’attivazione dei recettori M3 (Bell et al., 2013). Come negli studi di stimolazione elettrica, il tipo di risposta muscarinica non poteva essere correlato con i sottotipi anatomici di interneuroni. È importante notare che entrambi gli studi hanno dimostrato che gli interneuroni con proiezione perisomatica (probabilmente le cellule cestose che esprimono la parvalbumina) potrebbero rispondere al rilascio di acetilcolina con uno qualsiasi dei tre tipi di risposta muscarinica (Widmer et al., 2006; Bell et al., 2013). In diversi studi optogenetici, il potenziale di membrana degli interneuroni CA1 è stato valutato indirettamente misurando la frequenza sIPSC nei neuroni piramidali CA1 (Nagode et al., 2011, 2014). L’acetilcolina rilasciata optogeneticamente ha provocato un aumento di sIPSC di grande ampiezza con frequenze che rientravano nella larghezza di banda theta (4-12 Hz) (Nagode et al., 2011). È importante notare che questo aumento di sIPSCs potrebbe essere inibito dagli endocannabinoidi, suggerendo che essi derivano dall’attivazione di interneuroni positivi alla colecistochinina (Nagode et al., 2011). Inoltre, le sIPSCs non sono state influenzate dalla soppressione optogenetica delle cellule positive alla parvalbumina, suggerendo che non derivano dall’attivazione delle cellule cestose della parvalbumina, degli interneuroni axo-axonici, bistratificati o oriens-lacunosum-moleculare (Nagode et al., 2014). Questi risultati sono coerenti con gli studi di stimolazione sinaptica, che hanno registrato da un interneurone con morfologia di cellule cestose di colecistochinina che ha prodotto una risposta bifasica al rilascio di acetilcolina (Widmer et al., 2006). Quindi, basandosi solo sugli effetti sul potenziale di membrana, i recettori muscarinici attivati endogenamente sugli interneuroni CA1 ippocampali avranno effetti complessi sulla funzione della rete (vedi Tabella 1).
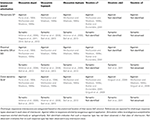
Tabella 1. Le risposte colinergiche variano in sottotipi di interneuroni anatomici simili e diversi.
Anche se diversi tipi di risposta muscarinica sono stati osservati quasi uniformemente in interneuroni CA1, non tutti i tipi di risposta sono stati evocati così facilmente dalla stimolazione optogenetica (Bell et al., 2013). Coerentemente con alcune registrazioni in vivo (Brazhnik e Fox, 1999), l’acetilcolina rilasciata dai terminali colinergici MS/DBB da lampi di luce blu consegnati a 20 Hz era in grado di produrre ogni tipo di risposta negli interneuroni CA1 ippocampali (Bell et al., 2013). Tuttavia, il numero di lampi ha influenzato la probabilità di osservare un particolare tipo di risposta. Negli interneuroni iperpolarizzanti, 10 flash erano sufficienti (91% degli interneuroni iperpolarizzanti) per osservare una risposta. Al contrario, 10 flash non erano sufficienti a produrre una risposta nella maggior parte degli interneuroni depolarizzanti (58%). Allo stesso modo, la fase depolarizzante non poteva essere osservata nella maggior parte degli interneuroni bifasici (55%) quando solo 10 stimoli sono stati consegnati. Pertanto, iperpolarizzazioni muscariniche possono richiedere meno attività colinergica presinaptica MS/DBB rispetto alle risposte depolarizzanti in interneuroni ippocampali CA1. Può essere che la soppressione dell’eccitabilità degli interneuroni sarà l’effetto predominante in risposta a bassi livelli di attività colinergica MS/DBB.
Effetti dell’attivazione dei recettori nicotinici sugli interneuroni ippocampali CA1
L’attivazione dei recettori nicotinici nell’ippocampo ha un impatto significativo sulla formazione della memoria fisiologica e fisiopatologica (Levin, 2002; Levin et al, 2002, 2009; Buccafusco et al., 2005; Davis e Gould, 2006, 2009; Nott e Levin, 2006; Davis et al., 2007). Delle 11 diverse subunità dei recettori nicotinici che si trovano nel SNC dei mammiferi, 9 sono state riportate per essere espresse nei neuroni ippocampali CA1 (Sudweeks e Yakel, 2000). Usando l’applicazione esogena di agonisti nicotinici, i recettori nicotinici funzionali che contengono α7 (Alkondon et al., 1997; Jones e Yakel, 1997; Frazier et al, 1998b; McQuiston e Madison, 1999c), α4β2 (McQuiston e Madison, 1999c; Sudweeks e Yakel, 2000), o α2 subunità (McQuiston e Madison, 1999c; Sudweeks e Yakel, 2000; Jia et al., 2009) sono stati osservati in interneuroni ippocampali CA1. Anche se gli interneuroni ippocampali sembravano esprimere una collezione diversificata di sottotipi di recettori nicotinici, i recettori contenenti α7 erano più frequentemente osservati e producevano risposte più grandi (McQuiston e Madison, 1999c; Sudweeks e Yakel, 2000). Infatti, i recettori nicotinici α7 nell’ippocampo sono stati associati alla formazione della memoria (Levin, 2002; Levin et al., 2002; Nott e Levin, 2006) e la loro disfunzione può avere un ruolo in alcune forme di schizofrenia (Freedman et al., 1994; Leonard et al., 1996; Adler et al., 1998). Tuttavia, nonostante i loro livelli di espressione più bassi, i recettori nicotinici contenenti α4β2 sono stati segnalati per svolgere un ruolo significativo nella formazione della memoria (Davis e Gould, 2006; Davis et al., 2007) e nella dipendenza da nicotina ippocampale-dipendente (Perry et al, 1999; Davis e Gould, 2009). I recettori contenenti α4β2 sono stati anche correlati ai deficit cognitivi associati all’invecchiamento e alla malattia di Alzheimer (Kellar et al., 1987; Wu et al., 2004; Gahring et al., 2005). Per comprendere appieno il ruolo che le diverse subunità nicotiniche svolgono nell’ippocampo, si è iniziato a studiare l’effetto dell’acetilcolina rilasciata endogena sulle singole cellule ippocampali e sulla rete ippocampale.
Attivazione dei recettori nicotinici negli interneuroni ippocampali CA1 in seguito al rilascio di acetilcolina
È stato dimostrato che il rilascio di acetilcolina dai terminali colinergici MS/DBB nell’ippocampo CA1 attiva i recettori nicotinici sugli interneuroni (Alkondon et al, 1998; Frazier et al., 1998a; Stone, 2007). Correnti postsinaptiche eccitatorie nicotiniche (EPSCs) sono state osservate per la prima volta usando la stimolazione elettrica e il patch clamping di cellule intere in fette acute di cervello di ratto. Queste EPSC nicotiniche avevano una cinetica veloce ed erano bloccate da antagonisti del recettore nicotinico α7 (Alkondon et al., 1998; Frazier et al., 1998a), coerentemente con gli studi che applicavano agonisti del recettore nicotinico direttamente sui corpi cellulari degli interneuroni (Alkondon et al., 1997; Jones e Yakel, 1997; Frazier et al., 1998b; McQuiston e Madison, 1999c). Tuttavia, più recenti studi optogenetici in fette di cervello di topo non sono stati in grado di riprodurre queste osservazioni precedenti (Bell et al., 2011). Invece, l’acetilcolina rilasciata optogeneticamente ha attivato principalmente i recettori nicotinici che contenevano subunità α4β2. Inoltre, le risposte α4β2 erano per lo più sottosoglia e avevano cinetiche molto lente. Questi dati erano suggestivi del fatto che l’acetilcolina diffondeva una distanza significativa prima di legarsi ai recettori nicotinici contenenti α4β2 (McQuiston e Madison, 1999c; Bennett et al., 2012), coerentemente con il volume o la trasmissione non sinaptica (Vizi et al., 2010). Anche se queste piccole risposte nicotiniche potrebbero sommare temporalmente, la loro capacità di eccitare interneuroni era limitata attraverso l’inibizione muscarinica presinaptica. Poiché le risposte nicotiniche erano per lo più sottosoglia, la trasmissione nicotinica sugli interneuroni CA1 può essere principalmente di natura modulatoria. Gli studi optogenetici hanno anche esaminato le risposte nicotiniche usando l’imaging del colorante sensibile al voltaggio (VSD). I segnali nicotinici VSD sono stati completamente bloccati dall’antagonista del recettore α4β2 DHβE e sono stati trovati significativamente più grandi nella regione dendritica distale dei neuroni piramidali CA1, che si sovrappone agli ingressi dalla corteccia entorinale e dal nucleo reuniens del talamo (Bell et al., 2011). Importante, perché il VSD macchia tutti gli elementi del tessuto, i dati VSD suggeriscono che α4β2 contenente recettori nicotinici sono il recettore più prevalente che media le risposte nicotiniche depolarizzanti in mouse ippocampale CA1. In particolare, le risposte nicotiniche potrebbero essere prodotte da un singolo flash di luce (Bell et al., 2011) suggerendo che il rilascio di acetilcolina dai terminali colinergici MS/DBB può aiutare a reclutare interneuroni attraverso l’attivazione dei recettori nicotinici prima che siano influenzati dall’attivazione dei recettori muscarinici.
Effetti del rilascio di acetilcolina sulla funzione della rete ippocampale CA1 dal punto di vista del potenziale di membrana degli interneuroni
Perché i potenziali di membrana degli interneuroni inibitori CA1 possono essere modulati in modo diverso dall’attivazione dei recettori muscarinici e nicotinici dopo il rilascio di acetilcolina, l’effetto conseguente sulla funzione della rete è senza dubbio complesso. L’attivazione dei recettori muscarinici può provocare effetti diversi e opposti, anche all’interno dello stesso interneurone (vedi tabella 1). Purtroppo, la nostra comprensione di come ogni sottotipo di interneurone può essere influenzato dall’attivazione dei recettori muscarinici o nicotinici rimane incompleta. Tuttavia, il numero di stimoli richiesti per produrre ogni tipo di risposta variava in modo coerente. Le risposte nicotiniche sono state evocate più facilmente richiedendo il minor numero di stimoli (Bell et al., 2011) mentre le risposte muscariniche depolarizzanti sono state le più difficili da produrre richiedendo il maggior numero di stimoli (Bell et al., 2013). Pertanto, si può ipotizzare che bassi livelli di attività dei neuroni colinergici MS/DBB e concentrazioni più basse di acetilcolina extracellulare favoriscono l’attivazione dei recettori nicotinici o una iperpolarizzazione muscarinica in sottoinsiemi specifici di interneuroni CA1.
Perché l’iperpolarizzazione muscarinica degli interneuroni CA1 richiede meno attività colinergica presinaptica, la disinibizione (attivazione indiretta) delle cellule piramidali CA1 ippocampali può essere favorita durante bassi livelli di attività colinergica MS/DBB (Figura 1B). Inoltre, postulando che le risposte nicotiniche colpiscono preferenzialmente interneuroni che inibiscono selettivamente altri interneuroni (interneurone-selettivo o IS), l’attivazione del recettore nicotinico può anche portare alla disinibizione dei neuroni piramidali CA1 (Figura 1A). Insieme, bassi livelli di attività colinergica MS/DBB favorirebbe una disinibizione netta di ippocampo CA1 permettendo maggiore probabilità di uscita da CA1 neuroni piramidali. L’aumento dell’output da CA1 può risultare nella facilitazione del richiamo e del consolidamento della memoria in altre aree del SNC, come si pensa avvenga durante il sonno ad onde lente (Gais e Born, 2004; Hasselmo e McGaughy, 2004). Al contrario, livelli più elevati di attività dei neuroni colinergici MS/DBB accoppiati a maggiori concentrazioni extracellulari di acetilcolina recluteranno successivamente diversi sottoinsiemi di interneuroni che rispondono tramite depolarizzazioni muscariniche. Alcuni di questi interneuroni depolarizzanti possono imporre l’inibizione ritmica dei neuroni piramidali CA1 a frequenze theta (Nagode et al., 2011, 2014), un ritmo di rete osservato durante livelli più elevati di rilascio di acetilcolina (Zhang et al., 2010). Questo si tradurrebbe in inibizione dell’output dei neuroni piramidali CA1 ippocampali (in parte ritmico), mentre facilita l’integrazione sinaptica all’interno dei dendriti delle cellule piramidali CA1 ippocampali attraverso effetti colinergici sui recettori glutammatergici e la funzione dendritica (Figura 1C) (Tsubokawa e Ross, 1997; Tsubokawa, 2000; Fernandez De Sevilla e Buno, 2010; Giessel e Sabatini, 2010). Infatti, un tale ruolo dinamico per le concentrazioni di acetilcolina nell’apprendimento e nella formazione della memoria è stato precedentemente proposto (Hasselmo, 2006; Hasselmo e Giocomo, 2006; Giocomo e Hasselmo, 2007; Hasselmo e Sarter, 2011). In questo schema, concentrazioni più basse di acetilcolina permettono alle interazioni sinaptiche intra-ippocampali (collaterali di Schaffer) di dominare, aumentando così l’output ippocampale CA1 e il recupero della memoria, mentre concentrazioni più alte di acetilcolina favoriscono l’elaborazione degli input dall’esterno dell’ippocampo, permettendo la formazione transitoria dei ricordi nella CA1 ippocampale. Pertanto, l’effetto combinato del rilascio di acetilcolina sugli input glutammatergici e sulla funzione degli interneuroni può svolgere un ruolo importante nella regolazione della rete ippocampale CA1 per il richiamo o per la formazione di nuovi ricordi.

Figura 1. Ipotesi che gli ingressi colinergici MS/DBB sopprimono o attivano le reti interneuronali nell’ippocampo CA1 a seconda dell’attività dei neuroni colinergici. (A) Bassi livelli di attività colinergica MS/DBB attivare preferenzialmente sottoinsiemi di interneuroni attraverso l’attivazione dei recettori nicotinici. Noi postuliamo che gli interneuroni guidati dalla nicotina sono interneuroni selettivi (IS, giallo-attivazione) che inibiscono specificamente altri interneuroni (blu). Aumentare la loro attività si traduce in disinibizione dei neuroni piramidali (P, giallo-attivazione, e uscita aumentata). (B) Bassi livelli di attività colinergica MS/DBB anche iperpolarizzare sottoinsiemi di interneuroni attraverso l’attivazione dei recettori muscarinici (I, blu-soppressione) con conseguente disinibizione dei neuroni piramidali (P, giallo-attivazione, e una maggiore uscita). (C) L’aumento dell’attività dei neuroni colinergici fa sì che sottoinsiemi di interneuroni siano depolarizzati dall’attivazione dei recettori muscarinici (I, attivazione rossa e aumento dell’inibizione sinaptica) con conseguente soppressione dei neuroni piramidali (P, uscita soppressa in blu).
Conflict of Interest Statement
L’autore dichiara che la ricerca è stata condotta in assenza di relazioni commerciali o finanziarie che potrebbero essere interpretate come un potenziale conflitto di interessi.
Riconoscimento
Questo lavoro è stato sostenuto da sovvenzioni del NIMH/NIH 5R01MH094626 e 1R21MH103695.
Adler, L. E., Olincy, A., Waldo, M., Harris, J. G., Griffith, J., Stevens, K., et al. Schizofrenia, gating sensoriale e recettori nicotinici. Schizofrenia. Bull. 24, 189-202. doi: 10.1093/oxfordjournals.schbul.a033320
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Alkondon, M., Pereira, E. F., e Albuquerque, E. X. (1998). recettori nicotinici sensibili all’alfa-bungarotossina e alla metilcaconitina mediano la trasmissione sinaptica veloce negli interneuroni di fette di ippocampo di ratto. Brain Res. 810, 257-263. doi: 10.1016/S0006-8993(98)00880-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Alkondon, M., Pereira, E. F., Barbosa, C. T., and Albuquerque, E. X. (1997). L’attivazione neuronale del recettore nicotinico dell’acetilcolina modula il rilascio di acido gamma-aminobutirrico dai neuroni CA1 di fette di ippocampo di ratto. J. Pharmacol. Exp. Ther. 283, 1396-1411.
Pubmed Abstract | Pubmed Full Text
Atri, A., Sherman, S., Norman, K. A., Kirchhoff, B. A., Nicolas, M. M., Greicius, M. D., et al. Blocco dei recettori colinergici centrali compromette il nuovo apprendimento e aumenta l’interferenza proattiva in una parola di coppia-associato compito di memoria. Behav. Neurosci. 118, 223-236. doi: 10.1037/0735-7044.118.1.223
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Aznavour, N., Mechawar, N., e Descarries, L. (2002). Analisi comparativa dell’innervazione colinergica nell’ippocampo dorsale del topo adulto e del ratto: uno studio quantitativo immunocitochimico. Hippocampus 12, 206-217. doi: 10.1002/hipo.1108
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Barrenechea, C., Pedemonte, M., Nunez, A., e Garcia-Austt, E. (1995). Registrazioni intracellulari in vivo della banda mediale settale e diagonale dei neuroni di Broca: relazioni con il ritmo theta. Exp. Brain Res. 103, 31-40. doi: 10.1007/BF00241962
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Behrends, J. C., e Ten Bruggencate, G. (1993). Modulazione colinergica dell’inibizione sinaptica nell’ippocampo della cavia in vitro: eccitazione degli interneuroni GABAergici e inibizione del rilascio di GABA. J. Neurophysiol. 69, 626-629.
Pubmed Abstract | Pubmed Full Text
Bell, K. A., Shim, H., Chen, C. K., e McQuiston, A. R. (2011). Potenziali postsinaptici eccitatori nicotinici in interneuroni ippocampali CA1 sono prevalentemente mediati da recettori nicotinici che contengono alfa4 e beta2 subunità. Neuropharmacology 61, 1379-1388. doi: 10.1016/j.neuropharm.2011.08.024
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Bell, L. A., Bell, K. A., and McQuiston, A. R. (2013). Tipi di risposta muscarinica sinaptica in interneuroni ippocampali CA1 dipendono da diversi livelli di attività presinaptica e diversi sottotipi di recettori muscarinici. Neuropharmacology 73, 160-173. doi: 10.1016/j.neuropharm.2013.05.026
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Bennett, C., Arroyo, S., Berns, D., e Hestrin, S. (2012). Meccanismi che generano dual-component nicotinici EPSCs in interneuroni corticali. J. Neurosci. 32, 17287-17296. doi: 10.1523/JNEUROSCI.3565-12.2012
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Blake, M. G., Krawczyk, M. C., Baratti, C. M., e Boccia, M. M. (2014). Neurofarmacologia del consolidamento e del riconsolidamento della memoria: approfondimenti sui meccanismi colinergici centrali. J. Physiol. Paris. doi: 10.1016/j.jphysparis.2014.04.005.
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Blokland, A., Honig, W., e Raaijmakers, W. G. (1992). Effetti delle iniezioni di scopolamina intra-ippocampale in un compito ripetuto di acquisizione spaziale nel ratto. Psicofarmacologia (Berl). 109, 373-376. doi: 10.1007/BF02245886
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Brazhnik, E. S., e Fox, S. E. (1997). Registrazioni intracellulari da neuroni del setto mediale durante il ritmo theta ippocampale. Exp. Brain Res. 114, 442-453. doi: 10.1007/PL00005653
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Brazhnik, E. S., e Fox, S. E. (1999). Potenziali d’azione e relazioni con il ritmo theta dei neuroni del setto mediale in vivo. Exp. Brain Res. 127, 244-258. doi: 10.1007/s002210050794
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Buccafusco, J. J., Letchworth, S. R., Bencherif, M., e Lippiello, P. M. (2005). Miglioramento cognitivo di lunga durata con agonisti del recettore nicotinico: meccanismi di discordanza farmacocinetica-farmacodinamica. Trends Pharmacol. Sci. 26, 352-360. doi: 10.1016/j.tips.2005.05.007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Buhler, A. V., and Dunwiddie, T. V. (2001). Regolazione dell’attività degli interneuroni dello strato oriens dell’ippocampo da parte dei recettori nicotinici alfa7 dell’acetilcolina. Neuroscience 106, 55-67. doi: 10.1016/S0306-4522(01)00257-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Buzsaki, G. (2002). Oscillazioni Theta nell’ippocampo. Neuron 33, 325-340. doi: 10.1016/S0896-6273(02)00586-X
CrossRef Full Text
Buzsaki, G., e Wang, X. J. (2012). Meccanismi di oscillazioni gamma. Annu. Rev. Neurosci. 35, 203-225. doi: 10.1146/annurev-neuro-062111-150444
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Cea-Del Rio, C. A., Lawrence, J. J., Erdelyi, F., Szabo, G., e McBain, C. J. (2011). Modulazione colinergica amplifica le proprietà oscillatorie intrinseche di CA1 interneuroni ippocampali colecistochinina-positivi. J. Physiol. 589, 609-627. doi: 10.1113/jphysiol.2010.199422
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Cea-Del Rio, C. A., Lawrence, J. J., Tricoire, L., Erdelyi, F., Szabo, G., and McBain, C. J. (2010). L’espressione del recettore muscarinico M3 dell’acetilcolina conferisce una modulazione colinergica differenziale ai sottotipi neurochimicamente distinti delle cellule del cesto ippocampale. J. Neurosci. 30, 6011-6024. doi: 10.1523/JNEUROSCI.5040-09.2010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Cobb, S. R., and Davies, C. H. (2005). Modulazione colinergica di cellule e circuiti ippocampali. J. Physiol. 562, 81-88. doi: 10.1113/jphysiol.2004.076539
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Cole, A. E., and Nicoll, R. A. (1983). L’acetilcolina media un potenziale sinaptico lento nelle cellule piramidali dell’ippocampo. Scienza 221, 1299-1301. doi: 10.1126/science.6612345
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Davis, J. A., and Gould, T. J. (2006). Gli effetti di DHBE e MLA sull’aumento indotto dalla nicotina del condizionamento di paura contestuale nei topi C57BL/6. Psicofarmacologia (Berl). 184, 345-352. doi: 10.1007/s00213-005-0047-y
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Davis, J. A., e Gould, T. J. (2009). I nAChR ippocampali mediano i deficit di apprendimento legati all’astinenza da nicotina. Eur. Neuropsychopharmacol. 19, 551-561. doi: 10.1016/j.euroneuro.2009.02.003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Davis, J. A., Kenney, J. W., and Gould, T. J. (2007). Coinvolgimento del recettore nicotinico dell’acetilcolina alpha4beta2 dell’ippocampo nel rafforzamento dell’effetto della nicotina acuta sul condizionamento della paura contestuale. J. Neurosci. 27, 10870-10877. doi: 10.1523/JNEUROSCI.3242-07.2007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Deiana, S., Platt, B., and Riedel, G. (2011). Il sistema colinergico e l’apprendimento spaziale. Comportamento. Brain Res. 221, 389-411. doi: 10.1016/j.bbr.2010.11.036
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Dutar, P., Bassant, M. H., Senut, M. C., and Lamour, Y. (1995). La via septohippocampale: struttura e funzione di un sistema colinergico centrale. Physiol. Rev. 75, 393-427.
Pubmed Abstract | Pubmed Full Text
Easton, A., Douchamps, V., Eacott, M., e Lever, C. (2012). Un ruolo specifico per l’acetilcolina septohippocampale nella memoria? Neuropsychologia 50, 3156-3168. doi: 10.1016/j.neuropsychologia.2012.07.022
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Fernandez De Sevilla, D., e Buno, W. (2010). Il potenziamento muscarinico a lungo termine della trasmissione mediata dai recettori NMDA e AMPA nelle sinapsi collaterali di Schaffer si sviluppa attraverso diversi meccanismi intracellulari. J. Neurosci. 30, 11032-11042. doi: 10.1523/JNEUROSCI.1848-10.2010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Frazier, C. J., Buhler, A. V., Weiner, J. L., e Dunwiddie, T. V. (1998a). Potenziali sinaptici mediati tramite recettori nicotinici nicotinici sensibili all’alfa-bungarotossina in interneuroni ippocampali di ratto. J. Neurosci. 18, 8228-8235.
Pubmed Abstract | Pubmed Full Text
Frazier, C. J., Rollins, Y. D., Breese, C. R., Leonard, S., Freedman, R., and Dunwiddie, T. V. (1998b). L’acetilcolina attiva una corrente nicotinica sensibile all’alfa-bungarotossina negli interneuroni ippocampali di ratto, ma non nelle cellule piramidali. J. Neurosci. 18, 1187-1195.
Pubmed Abstract | Pubmed Full Text
Freedman, R., Adler, L. E., Bickford, P., Byerley, W., Coon, H., Cullum, C. M., et al. Schizofrenia e recettori nicotinici. Harv. Rev. Psychiatry 2, 179-192. doi: 10.3109/10673229409017136
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Freund, T. F., and Buzsaki, G. (1996). Interneuroni dell’ippocampo. Hippocampus 6, 347-470.
Pubmed Abstract | Pubmed Full Text
Fukudome, Y., Ohno-Shosaku, T., Matsui, M., Omori, Y., Fukaya, M., Tsubokawa, H., et al. Due classi distinte di azione muscarinica sulle sinapsi inibitorie ippocampali: M2-mediata soppressione diretta e M1/M3-mediata soppressione indiretta attraverso la segnalazione di endocannabinoidi. Eur. J. Neurosci. 19, 2682-2692. doi: 10.1111/j.0953-816X.2004.03384.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Gahring, L. C., Persiyanov, K., and Rogers, S. W. (2005). Cambiamenti specifici del ceppo del topo nell’espressione del recettore nicotinico con l’età. Neurobiol. Aging 26, 973-980. doi: 10.1016/j.neurobiolaging.2004.07.005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Gais, S., and Born, J. (2004). Bassa acetilcolina durante il sonno ad onde lente è fondamentale per il consolidamento della memoria dichiarativa. Proc. Natl. Acad. Sci. U.S.A. 101, 2140-2144. doi: 10.1073/pnas.0305404101
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Giessel, A. J., and Sabatini, B. L. (2010). I recettori muscarinici M1 aumentano i potenziali sinaptici e l’afflusso di calcio nelle spine dendritiche inibendo i canali SK postsinaptici. Neuron 68, 936-947. doi: 10.1016/j.neuron.2010.09.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Giocomo, L. M., and Hasselmo, M. E. (2007). La neuromodulazione da glutammato e acetilcolina può cambiare la dinamica del circuito regolando l’influenza relativa dell’input afferente e del feedback eccitatorio. Mol. Neurobiol. 36, 184-200. doi: 10.1007/s12035-007-0032-z
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Griguoli, M., Scuri, R., Ragozzino, D., e Cherubini, E. (2009). L’attivazione dei recettori nicotinici dell’acetilcolina aumenta una conduttanza lenta del potassio calcio-dipendente e riduce l’accensione degli interneuroni dello strato oriens. Eur. J. Neurosci. 30, 1011-1022. doi: 10.1111/j.1460-9568.2009.06914.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hasselmo, M. E. (2006). Il ruolo dell’acetilcolina nell’apprendimento e nella memoria. Curr. Opin. Neurobiol. 16, 710-715. doi: 10.1016/j.conb.2006.09.002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hasselmo, M. E., and Giocomo, L. M. (2006). Modulazione colinergica della funzione corticale. J. Mol. Neurosci. 30, 133-135. doi: 10.1385/JMN:30:1:133
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hasselmo, M. E., and McGaughy, J. (2004). Alti livelli di acetilcolina impostano le dinamiche del circuito per l’attenzione e la codifica e bassi livelli di acetilcolina impostano le dinamiche per il consolidamento. Prog. Brain Res. 145, 207-231. doi: 10.1016/S0079-6123(03)45015-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hasselmo, M. E., e Sarter, M. (2011). Modalità e modelli di neuromodulazione colinergica forebrain della cognizione. Neuropsychopharmacology 36, 52-73. doi: 10.1038/npp.2010.104
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Jia, Y., Yamazaki, Y., Nakauchi, S., and Sumikawa, K. (2009). Alfa2 recettori della nicotina funzione come un interruttore molecolare per eccitare continuamente un sottoinsieme di interneuroni nei circuiti ippocampali di ratto. Eur. J. Neurosci. 29, 1588-1603. doi: 10.1111/j.1460-9568.2009.06706.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Jones, S., and Yakel, J. L. (1997). Recettori nicotinici funzionali di ACh sugli interneuroni nell’ippocampo del ratto. J. Physiol. 504(Pt 3), 603-610. doi: 10.1111/j.1469-7793.1997.603bd.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Kellar, K. J., Whitehouse, P. J., Martino-Barrows, A. M., Marcus, K., and Price, D. L. (1987). Siti di legame colinergici muscarinici e nicotinici nella corteccia cerebrale della malattia di Alzheimer. Brain Res. 436, 62-68. doi: 10.1016/0006-8993(87)91556-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Kenney, J. W., and Gould, T. J. (2008). Modulazione dell’apprendimento ippocampo-dipendente e la plasticità sinaptica da nicotina. Mol. Neurobiol. 38, 101-121. doi: 10.1007/s12035-008-8037-9
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Klausberger, T., e Somogyi, P. (2008). Diversità neuronale e dinamica temporale: l’unità delle operazioni del circuito ippocampale. Scienza 321, 53-57. doi: 10.1126/science.1149381
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Larkum, M. E., Zhu, J. J., e Sakmann, B. (1999). Un nuovo meccanismo cellulare per l’accoppiamento degli input che arrivano a diversi strati corticali. Nature 398, 338-341. doi: 10.1038/18686
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Lawrence, J. J., Statland, J. M., Grinspan, Z. M., e McBain, C. J. (2006). Tipo di cellula-specifica dipendenza di segnalazione muscarinica in ippocampo mouse stratum oriens interneuroni. J. Physiol. 570, 595-610. doi: 10.1113/jphysiol.2005.100875
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Leonard, S., Adams, C., Breese, C. R., Adler, L. E., Bickford, P., Byerley, W., et al. Funzione dei recettori nicotinici nella schizofrenia. Schizophr. Bull. 22, 431-445. doi: 10.1093/schbul/22.3.431
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Levin, E. D. (2002). Sottotipi di recettori nicotinici e funzione cognitiva. J. Neurobiol. 53, 633-640. doi: 10.1002/neu.10151
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Levin, E. D., Bradley, A., Addy, N., and Sigurani, N. (2002). Recettori nicotinici ippocampali alfa 7 e alfa 4 beta 2 e memoria di lavoro. Neuroscience 109, 757-765. doi: 10.1016/S0306-4522(01)00538-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Levin, E. D., Petro, A., Rezvani, A. H., Pollard, N., Christopher, N. C., Strauss, M., et al. Knotinic alfa7- o beta2-containing receptor knockout: effetti sull’apprendimento del labirinto a braccia radiali e sul consumo a lungo termine di nicotina nei topi. Comportamento. Brain Res. 196, 207-213. doi: 10.1016/j.bbr.2008.08.048
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
McQuiston, A. R., and Madison, D. V. (1999a). L’attività dei recettori muscarinici ha effetti multipli sui potenziali di membrana a riposo degli interneuroni ippocampali CA1. J. Neurosci. 19, 5693-5702.
Pubmed Abstract | Pubmed Full Text
McQuiston, A. R., and Madison, D. V. (1999b). L’attività dei recettori muscarinici induce una postdepolarizzazione in una sottopopolazione di interneuroni CA1 ippocampali. J. Neurosci. 19, 5703-5710.
Pubmed Abstract | Pubmed Full Text
McQuiston, A. R., and Madison, D. V. (1999c). L’attivazione dei recettori nicotinici eccita sottotipi distinti di interneuroni nell’ippocampo di ratto. J. Neurosci. 19, 2887-2896.
Pubmed Abstract | Pubmed Full Text
Miles, R., Toth, K., Gulyas, A. I., Hajos, N., and Freund, T. F. (1996). Differenze tra inibizione somatica e dendritica nell’ippocampo. Neuron 16, 815-823. doi: 10.1016/S0896-6273(00)80101-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Nagode, D. A., Tang, A. H., Karson, M. A., Klugmann, M., e Alger, B. E. (2011). Rilascio optogenetica di ACh induce raffiche ritmiche di IPSCs perisomatico in ippocampo. PLoS ONE 6:e27691. doi: 10.1371/journal.pone.0027691
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Nagode, D. A., Tang, A. H., Yang, K., e Alger, B. E. (2014). Identificazione optogenetica di un intrinseco colinergicamente guidato oscillatore inibitorio sensibile ai cannabinoidi e oppioidi in ippocampo CA1. J. Physiol. 592, 103-123. doi: 10.1113/jphysiol.2013.257428
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Nott, A., and Levin, E. D. (2006). Dorsale ippocampale alfa7 e alfa4beta2 recettori nicotinici e la memoria. Brain Res. 1081, 72-78. doi: 10.1016/j.brainres.2006.01.052
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Parra, P., Gulyas, A. I., and Miles, R. (1998). Quanti sottotipi di cellule inibitorie nell’ippocampo? Neuron 20, 983-993. doi: 10.1016/S0896-6273(00)80479-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Perry, D. C., Davila-Garcia, M. I., Stockmeier, C. A., and Kellar, K. J. (1999). Aumento dei recettori nicotinici nel cervello dei fumatori: studi di legame di membrana e autoradiografia. J. Pharmacol. Exp. Ther. 289, 1545-1552.
Pubmed Abstract | Pubmed Full Text
Pitler, T. A., and Alger, B. E. (1992). Eccitazione colinergica di interneuroni GABAergici nella fetta ippocampale di ratto. J. Physiol. 450, 127-142.
Pubmed Abstract | Pubmed Full Text
Potenza, A. E., Vazdarjanova, A., e McGaugh, J. L. (2003). Influenze colinergiche muscariniche nel consolidamento della memoria. Neurobiol. Imparare. Mem. 80, 178-193. doi: 10.1016/S1074-7427(03)00086-8
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Simon, A. P., Poindessous-Jazat, F., Dutar, P., Epelbaum, J., e Bassant, M. H. (2006). Proprietà di cottura dei neuroni anatomicamente identificati nel setto mediale di ratti anestetizzati e non anestetizzati trattenuti. J. Neurosci. 26, 9038-9046. doi: 10.1523/JNEUROSCI.1401-06.2006
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Smiley, J. F., Morrell, F., and Mesulam, M. M. (1997). Sinapsi colinergiche nella corteccia cerebrale umana: uno studio ultrastrutturale in sezioni seriali. Exp. Neurol. 144, 361-368. doi: 10.1006/exnr.1997.6413
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Stone, T. W. (2007). L’acido kynurenico blocca la trasmissione sinaptica nicotinica agli interneuroni ippocampali nei giovani ratti. Eur. J. Neurosci. 25, 2656-2665. doi: 10.1111/j.1460-9568.2007.05540.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Storm-Mathisen, J. (1970). Istochimica quantitativa di acetilcolinesterasi nella regione ippocampale di ratto correlata alla colorazione istochimica. J. Neurochem. 17, 739-750. doi: 10.1111/j.1471-4159.1970.tb03344.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Sudweeks, S. N., and Yakel, J. L. (2000). Caratterizzazione funzionale e molecolare dei recettori nicotinici ACh neuronali nei neuroni ippocampali CA1 del ratto. J. Physiol. 527(Pt 3), 515-528. doi: 10.1111/j.1469-7793.2000.00515.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Szabo, G. G., Holderith, N., Gulyas, A. I., Freund, T. F., and Hajos, N. (2010). Proprietà sinaptiche distinte dei tipi di cellule inibitorie perisomatiche e la loro diversa modulazione dall’attivazione del recettore colinergico nella regione CA3 dell’ippocampo del topo. Eur. J. Neurosci. 31, 2234-2246. doi: 10.1111/j.1460-9568.2010.07292.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Tang, A. H., Karson, M. A., Nagode, D. A., McIntosh, J. M., Uebele, V. N., Renger, J. J., et al. I recettori nicotinici terminali nervosi dell’acetilcolina iniziano il rilascio quantico di GABA dagli interneuroni perisomatici attivando i canali assonali T-type (Cav3) Ca(2)(+) e il rilascio di Ca(2)(+) dai depositi. J. Neurosci. 31, 13546-13561. doi: 10.1523/JNEUROSCI.2781-11.2011
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Teles-Grilo Ruivo, L. M., e Mellor, J. R. (2013). Modulazione colinergica della funzione di rete ippocampale. Fronte. Neurosci sinaptico. 5:2. doi: 10.3389/fnsyn.2013.00002
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Tsubokawa, H. (2000). Controllo del Na+ spike backpropagation da segnalazione intracellulare nei dendriti del neurone piramidale. Mol. Neurobiol. 22, 129-141. doi: 10.1385/MN:22:1-3:129
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Tsubokawa, H., e Ross, W. N. (1997). Modulazione muscarinica della retropropagazione degli spike nei dendriti apicali dei neuroni piramidali CA1 dell’ippocampo. J. Neurosci. 17, 5782-5791.
Pubmed Abstract | Pubmed Full Text
Turrini, P., Casu, M. A., Wong, T. P., De, K. Y., Ribeiro-Da-Silva, A., and Cuello, A. C. (2001). I terminali nervosi colinergici stabiliscono sinapsi classiche nella corteccia cerebrale del ratto: modello sinaptico e atrofia legata all’età. Neuroscience 105, 277-285. doi: 10.1016/S0306-4522(01)00172-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Umbriaco, D., Garcia, S., Beaulieu, C., and Descarries, L. (1995). Caratteristiche relazionali dei terminali assonali di acetilcolina, noradrenalina, serotonina e GABA nello strato radiato dell’ippocampo di ratto adulto (CA1). Hippocampus 5, 605-620. doi: 10.1002/hipo.450050611
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Umbriaco, D., Watkins, K. C., Descarries, L., Cozzari, C., and Hartman, B. K. (1994). Caratteristiche ultrastrutturali e morfometriche dell’innervazione dell’acetilcolina nella corteccia parietale del ratto adulto: uno studio al microscopio elettronico in sezioni seriali. J. Comp. Neurol. 348, 351-373. doi: 10.1002/cne.903480304
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Vizi, E. S., Fekete, A., Karoly, R., e Mike, A. (2010). Recettori non sinaptici e trasportatori coinvolti nelle funzioni cerebrali e obiettivi del trattamento farmacologico. Br. J. Pharmacol. 160, 785-809. doi: 10.1111/j.1476-5381.2009.00624.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Widmer, H., Ferrigan, L., Davies, C. H., and Cobb, S. R. (2006). Potenziali sinaptici muscarinici acetilcolinergici lenti evocati in interneuroni ippocampali di ratto. Hippocampus 16, 617-628. doi: 10.1002/hipo.20191
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Wu, J., Kuo, Y. P., George, A. A, Xu, L., Hu, J., e Lukas, R. J. (2004). La beta-amiloide inibisce direttamente i recettori umani alfa4beta2-nicotinici dell’acetilcolina espressi eterologicamente in cellule umane SH-EP1. J. Biol. Chem. 279, 37842-37851. doi: 10.1074/jbc.M400335200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Zhang, H., Lin, S. C., e Nicolelis, M. A. (2010). Accoppiamento spazio-temporale tra il rilascio di acetilcolina ippocampale e oscillazioni theta in vivo. J. Neurosci. 30, 13431-13440. doi: 10.1523/JNEUROSCI.1144-10.2010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text