Per capire meglio questo meccanismo, una semplice ripartizione della via dell’odore è fornita qui sotto. Quando gli esseri umani masticano, i composti aromatici volatili sono spinti attraverso il rinofaringe e i recettori dell’olfatto.
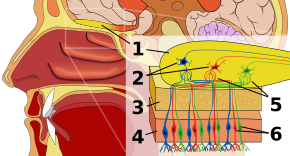
Epitelio olfattivoModifica
La prima tappa del sistema olfattivo è l’epitelio olfattivo, o tessuto appoggiato sul tetto della cavità nasale che ospita i recettori dell’odore. I recettori olfattivi sono neuroni bipolari che legano gli odori dell’aria e si riuniscono al nervo olfattivo prima di passare gli assoni ai dendriti delle cellule mitrali nel bulbo olfattivo. I recettori sensoriali nella bocca e nel naso sono polarizzati allo stato di riposo, e si depolarizzano in risposta a qualche cambiamento nell’ambiente, come il contatto con molecole di odore. Le molecole di odore, costituite da catene di idrocarburi con gruppi funzionali, si legano ai recettori sensoriali nel naso e nella bocca. Le proprietà dei gruppi funzionali includono: (1) lunghezza della catena di carbonio, (2) gruppo terminale, che concorda con le differenze associate ai diversi odori, (3) gruppo laterale, (4) chiralità, (5) forma e (6) dimensione. Quando le molecole di odore si legano ai recettori sensoriali, lo fanno in base a queste proprietà. Ogni cellula olfattiva ha un singolo tipo di recettore, ma quel recettore può essere “ampiamente sintonizzato” e le molecole di odore interagiscono ulteriormente a livello del recettore, nel senso che, in alcuni casi, una molecola di odore da sola potrebbe non legarsi a un recettore, ma in presenza di un’altra molecola di odore, l’originale si legherebbe e quindi creerebbe una sensazione di odore solo in presenza della seconda molecola.
Bulbo olfattivoModifica
Nel bulbo olfattivo, le molecole di odore sono mappate spazialmente. Queste rappresentazioni spaziali sono note come “immagini olfattive”. La rappresentazione spaziale permette l’inibizione laterale, o l’aumento del contrasto e la compressione del guadagno. L’aumento del contrasto è sensibile al cambiamento e mette in evidenza gli stimoli nel cervello che stanno cambiando piuttosto che a riposo. La compressione del guadagno aumenta la sensibilità agli stimoli a bassa intensità mentre diminuisce la sensibilità agli stimoli ad alta intensità. Il bulbo olfattivo, anche se ancora nelle fasi primarie della sua comprensione da parte dei ricercatori, distingue l’odore dagli altri sensi perché segna una deviazione nel percorso sensoriale da ciò che è caratteristico di tutti gli altri sensi. Vale a dire, tutte le informazioni sensoriali non olfattive passano attraverso il talamo dopo il livello del recettore, ma il fatto che le informazioni dell’odore invece entrino nella propria area specializzata potrebbe suggerire la storia primitiva dell’odore e/o un tipo distinto di elaborazione delle informazioni dell’odore sulla sua strada verso la corteccia. Il bulbo olfattivo ospita glomeruli, o giunzioni cellulari, su cui convergono migliaia di recettori dello stesso tipo, oltre alle cellule mitrali. Questa organizzazione permette di rappresentare concisamente una grande quantità di informazioni senza richiedere un numero altrettanto grande di tipi di recettori. La combinazione risultante di informazioni di odore è soprannominata un’immagine di odore a livello del bulbo olfattivo.
Imaging nel bulbo olfattivoModifica
Metodo 2DGModifica
Nel 1977, il biochimico Lou Sokoloff, Seymour Kety, e Floyd E. Bloom hanno sviluppato un modo di mappare l’attività del cervello tracciando la metabolizzazione dell’ossigeno nel cervello del ratto. Le cellule nervose richiedono ossigeno e glucosio per l’energia. Il 2-deossiglucosio (2DG) è un isotopo radioattivo del glucosio che può essere tracciato nel cervello poiché lascia una traccia nella cellula dove verrebbe normalmente metabolizzato per l’energia se fosse glucosio. Dopo la stimolazione di una certa regione di cellule, le fotografie a raggi X possono essere affettate per rivelare quali cellule erano attive, in particolare nelle giunzioni sinaptiche.
La risonanza magnetica funzionale (fMRI) può anche essere usata per misurare il metabolismo di un odore. Questo metodo non è terminale come il metodo del 2-deossiglucosio, così un animale può essere misurato con molti odori, e le immagini risultanti possono essere confrontate.
Metodo della proteina fluorescente verdeModifica
Infine, il metodo della proteina fluorescente verde ingegnerizza geneticamente i topi per esprimere una proteina nei neuroni attivi, e una telecamera può quindi essere posizionata all’interno del cranio del topo per misurare l’attività.
RisultatiModifica
Questi metodi rivelano, in particolare, che l’organizzazione delle informazioni olfattive nel bulbo olfattivo è spaziale. Modelli molecolari simili risultano in modelli di attivazione simili per quanto riguarda i glomeruli, e i glomeruli che sono più vicini tra loro codificano caratteristiche simili delle informazioni olfattive.
Corteccia olfattivaModifica
La corteccia olfattiva a tre strati, contenente cellule piramidali è il prossimo punto di riferimento sulla via dell’odore. Una cellula piramidale riceve informazioni da una molteplicità di cellule mitrali dal bulbo olfattivo, rendendo il modello glomerulare precedentemente organizzato distribuito nella corteccia olfattiva. Questa dispersione delle informazioni delle cellule mitrali permette connessioni di feedback autoeccitatorie, eccitazione laterale e auto e inibizione laterale. Questi processi contribuiscono all’apprendimento hebbiano, che prende il nome da Donald O. Hebb, ed è spesso semplificato dal detto “i neuroni che sparano insieme legano insieme”. Il potenziamento a lungo termine, il meccanismo neurale dell’apprendimento hebbiano, permette la formazione della memoria a livello delle cellule piramidali. L’apprendimento hebbiano è quindi essenzialmente il fenomeno per cui la corteccia olfattiva “ricorda” l’output delle combinazioni di molecole olfattive e permette il riconoscimento delle combinazioni precedentemente percepite più velocemente di quelle nuove, abbinandole agli input memorizzati. Gli odori risultanti, che prima erano chiamati immagini odorose, vengono memorizzati nella corteccia olfattiva per il riconoscimento e ora vengono chiamati oggetti odorosi. L’esperienza rafforza quindi il rapporto segnale-rumore in quanto un oggetto odoroso precedentemente percepito può essere più facilmente distinto contro un maggiore rumore di fondo.
Corteccia orbitofrontaleModifica
La corteccia orbitofrontale (OFC) è la destinazione finale delle informazioni odorose ed è dove nasce la percezione cosciente dell’odore. Le informazioni olfattive entrano direttamente dopo essere passate attraverso la corteccia olfattiva, che segna la distinzione dalle altre informazioni sensoriali che passano prima attraverso il talamo. L’OFC si trova dorsalmente alla corteccia prefrontale, permettendo alle informazioni olfattive un input diretto alla corteccia prefrontale, o la principale area decisionale del cervello. Ci sono solo tre tipi di neuroni che le molecole olfattive attraversano prima di arrivare all’OFC: le cellule recettrici olfattive nell’epitelio olfattivo, le cellule mitrali e i neuroni piramidali olfattivi.
A livello dell’OFC, vengono fatte associazioni con altre aree cerebrali, compresi gli input dalla bocca (somatosensazione), input emotivi (amigdala), informazioni visive e di valutazione (corteccia prefrontale). L’OFC è responsabile della sintonizzazione selettiva degli odori, della fusione dei domini sensoriali e delle valutazioni edonistiche degli odori.