前章で簡単に説明したように、神経伝達物質はシナプス前神経末端からシナプス間隙に放出される化学信号のことです。 その後、神経伝達物質がシナプス後神経細胞(または他のクラスの標的細胞)上の特定の受容体に結合すると、標的細胞の電気的特性が一過性に変化し、非常に多様なシナプス後効果をもたらす(第7章および第8章参照)。
電気情報が化学シグナルによってあるニューロンから次のニューロンに伝達できるという概念は、20世紀の前半を通じて激しい議論の対象であった。 この考えを裏付ける重要な実験が、1926年にドイツの生理学者オットー・ローウィによって行われました。 迷走神経を電気的に刺激すると、化学的な信号が放出され、心拍が遅くなることを証明したのである。 彼は、2匹のカエルの心臓を分離して灌流し、その拍動速度をモニターした(図6.1)。 彼の実験の要点は、刺激を受けた心臓を流れる灌流液を集め、それを2番目の心臓に移すことであった。 迷走神経は、灌流液中に蓄積される化学物質を放出することによって、心拍数を調節していることが明らかになった。 当初は「迷走神経物質」と呼ばれていたこの物質は、後にアセチルコリン(ACh)であることが判明し、長年にわたって最も徹底的に研究されてきた神経伝達物質となった。 AChは心臓だけでなく、中枢神経系や末梢神経系のさまざまなシナプス後標的で作用し、特に線条筋肉の神経筋接合部や内臓運動系で顕著である(第5、21章参照)
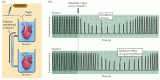
Figure 6.1
Loewi’s experiment demonstrating chemical neurotransmission. (A)実験のセットアップの図。 (B)分離したカエルの心臓の迷走神経を刺激したところ、心拍数が減少した(上図)。 刺激された心臓からの灌流液が、(さらに…)
長年の間に、ある物質を神経伝達物質と決定的に特定する正式な基準がいくつか出現している(Box A)。 それにもかかわらず、特定のシナプスで活性化する神経伝達物質を同定することは依然として困難であり、多くのシナプス(特に脳)において、神経伝達物質の性質は十分に確立されていない。 ボックスAで概説した基準をすべて満たしていない物質は、「推定」神経伝達物質と呼ばれる。
Box A
Criteria That Define a Neurotransmitter.comでは、神経伝達物質を定義する基準として、ボックスAで概説した基準すべてを満たしていない物質が、「推定」神経伝達物質として参照される。
他のシグナル伝達分子と比較して、神経伝達物質の際立った特徴は、内分泌系から分泌されるホルモンの作用と比較することで明確になる。 ホルモンは通常、ホルモンを分泌する細胞から遠く離れた標的細胞に影響を与える(第8章参照)。 この「離れた場所での作用」は、ホルモンが血流に放出されることで実現される。 これに対して、神経伝達物質が作用する距離はごくわずかである。 多くのシナプスでは、伝達物質はシナプス前末端の直下にあるシナプス後細胞の受容体にのみ結合する(図6.2A)。このような場合、伝達物質が作用する距離はマイクロメートル未満である。 神経伝達物質が局所的に拡散して、近傍の複数のシナプス後細胞(時にはシナプス前細胞)の電気的特性を変化させる場合でさえ(図6.2B)、数十から数百マイクロメートルの距離にわたってしか作用しないのである。 神経細胞の軸索突起が細長いため、神経細胞本体から1メートルも離れた場所で神経伝達物質が放出されることがあるが、これらの伝達物質は依然として放出されたシナプス前部位付近でのみ作用する(図6.2C)

図6.2
神経伝達物質作用の局在化。 神経伝達物質は一般に、単一のシナプス後細胞の小さな領域の電気的興奮性を変化させることによって局所的に(A)、またはより拡散的に(B)、(より…)電気的興奮性を変化させることによって作用する
神経伝達物質とホルモンの区別は概して明確であるが、物質は脳のある領域では神経伝達物質として作用しながら他の場所ではホルモンとして機能しうる。 例えば、バソプレシンやオキシトシンというペプチドホルモンは、下垂体後葉から循環器内に放出されますが、中枢のシナプスでは神経伝達物質としても機能しています。 その他にも、ホルモンと神経伝達物質の両方の働きをするペプチドが多数ある
。