- Introduction
- Results
- Cell Accumulation and Attachment to Mineral Requires Chemotaxis and Extracellular Electron Transport Genes in S. oneidensis MR-1
- Characterizing Congregation rond IEA in andere Shewanella stammen
- Discussie
- Materialen en methoden
- Kweek en stammen
- Minerale Synthese
- Cell Accumulation after Photo-Bleaching (CAAP) Time-Lapse Experiments
- Microscopie Capture van celbewegingen
- Zwemmen Analysis
- Author Contributions
- Conflict of Interest Statement
- Acknowledgments
- Aanvullend materiaal
Introduction
Aan het eind van de jaren tachtig werd aangetoond dat Shewanella oneidensis MR-1 (Myers en Nealson, 1988a) en later verschillende soorten Geobacter (Lovley et al., 1993; Champine et al., 2000) bleken in staat te zijn tot elektronenoverdracht naar onoplosbare elektronenacceptoren (IEA’s), zoals onoplosbare metaaloxiden en/of geladen elektroden: een proces dat extracellulair elektronentransport (EET) wordt genoemd (Myers en Nealson, 1988b; Venkateswaran et al., 1999; Bond en Lovley, 2003). Dit vermogen wekte aanzienlijke belangstelling met betrekking tot biogeochemische kringlopen, bioremediëring, corrosie, verwerking van nanomaterialen en energieproductie (Bretschger et al., 2007; Kan et al., 2011; Hsu et al., 2012). Hoewel bekend is dat verschillende groepen microben in staat zijn tot EET, zijn belangrijke mechanistische studies verricht met slechts twee modelsystemen, Shewanella (Fredrickson et al., 2008; Shi et al., 2009), en Geobacter (Lovley et al., Tot nu toe zijn meer dan 100 andere stammen van Shewanella geïsoleerd uit een grote verscheidenheid van habitats, waaronder open waterkolom, zandsteenschalie, mariene en zoetwatersedimenten, oliepijpleidingen, oliepekel, en zelfs algengemeenschappen boven op Antarctisch ijs (Hau en Gralnick, 2007). De genomen van meer dan 20 van deze soorten zijn volledig gesequenced (Fredrickson et al., 2008). Van verschillende van deze soorten is aangetoond dat ze in staat zijn tot EET tot IEA, waaronder S. oneidensis MR-1, Shewanella putrefaciens CN32, S. sp. ANA-3, S. sp. W3-18-1, Shewanella amazonensis SB2B, Shewanella frigidimarina NCIMB 400, en Shewanella loihica PV-4 (Fredrickson et al., 1998; Venkateswaran et al., 1998; Gao et al., 2006; Bretschger, 2008). Andere leden van het Shewanella-geslacht, zoals Shewanella baltica OS217 en Shewanella denitrificans OS155 (tabel 1) zijn niet in staat tot EET (Brettar et al., 2002). Van al deze stammen is de congregatie in reactie op IEA alleen bestudeerd voor MR-1.

Tabel 1. Genetische vergelijking van Shewanella spp. en hun oorspronkelijke habitat.
In het genus Shewanella is het EET-mechanisme van MR-1 het meest uitvoerig gekarakteriseerd. MR-1 maakt gebruik van verschillende benaderingen voor de reductie van onoplosbare IEA’s: (1) directe EET via extracellulaire meerkleurige cytochromen (Beliaev en Saffarini, 1998; Myers en Myers, 2001, 2002; Meyer et al., 2004; Mitchell et al., 2012; Kracke et al., 2015) (Figuur 1A); (2) gemedieerde EET met behulp van oplosbare elektronenshuttles gebonden aan membraancytochromen (Lovley et al., 1996; Marsili et al., 2008; Li et al., 2012; Kotloski and Gralnick, 2013; Okamoto et al, 2014); (3) gemedieerde EET door gebruik te maken van geleidende buitenmembraanuitlopers die cytochromen bevatten (Gorby et al., 2006; El-Naggar et al., 2010); en (4) geleidende extracellulaire matrices die geleidende en halfgeleidende mineralen bevatten (Kato et al., 2010). In stam MR-1 zijn verschillende genen geïdentificeerd waarvan is aangetoond dat ze essentieel zijn voor EET (figuur 1A en tabel 2), waaronder het tetraheme cytochroom c cymA (SO_4591) en de combinatie mtrBC/omcA (SO_1776, SO_1778 en SO_1779) die coderen voor de decaheme cytochroom c-component en het tetraheme cytochroom c dat nodig is voor de reductie van verschillende anaerobe elektronenacceptoren, waaronder metaaloxiden (Myers en Myers, 2001, 2002; Schwalb et al., 2003). Omdat al deze mechanismen vertrouwen op de cel nabijheid van IEA voor EET, is het belangrijk om te begrijpen de cel sensing en netto zwemmen migratie in de richting van de IEA. Met betrekking tot de gemeente, redox taxis of energie taxis, zijn veel studies uitgevoerd op MR-1 als gevolg van de veelzijdige elektronen acceptor gebruik (Bencharit en Ward, 2005; Baraquet et al., 2009; Harris et al., 2010). Energietaxis is een term die in ruime zin aerotaxis, fototaxis, redoxtaxis, taxis naar alternatieve elektronenacceptoren en chemotaxis naar oxideerbare substraten omvat (Alexandre et al., 2004).
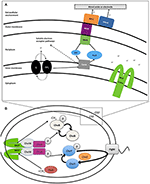
Figuur 1. (A) Extracellulair elektronentransport (EET) in Shewanella oneidensis MR-1 bouwt proton motive force (pmf). Onder anaërobe omstandigheden kunnen leden van het genus Shewanella elektronen transporteren vanuit het binnenmembraan, het periplasma, het buitenmembraan en een elektrode via een keten van cytochromen en menaquinonen (MQ) die gezamenlijk bekend staan als extracellulair elektronentransport (EET). De expressie van EET-cytochromen kan fluctueren afhankelijk van de in de omgeving beschikbare elektronenacceptor. Het aantal ijzerhoudende c-type heemgroepen is aangegeven binnen (haakjes). Elektronen stromen van elektronendonoren, zoals lactaat, naar de reductie van chinonen (Q), die tegelijkertijd H+ overdragen naar het periplasma, waardoor pmf wordt opgebouwd, terwijl ook elektronen worden doorgegeven aan CymA. Bij afwezigheid van oplosbare elektronenacceptoren worden de elektronen overgedragen aan het buitenmembraancomplex MtrCAB. Dit MtrCAB-complex kan elektronen rechtstreeks afstaan aan de eindelektronenacceptor, hetzij mineraal of anode-elektrode, hetzij via flavinmoleculen. De pmf drijft de productie van ATP en de rotatie van de polaire flagella aan. “Self-sensing” methyl-accepterende chemotaxis-eiwitten (MCPs) controleren de rotatie van de flagella via het chemotaxis-signaaltransductiesysteem (B) en kunnen veranderingen in de H+-concentratie tijdens metaalreductie detecteren. (B) Waarschijnlijke chemotaxis signaal transductie pathway in Shewanella. Als reactie op stimulatie verschuift de structuur van MCP als een zuiger, waardoor de autofosforylering van CheA wordt vertraagd of gestopt. CheY en CheB worden daardoor niet gefosforyleerd, en dit gebrek aan CheY-P maakt soepel zwemmen mogelijk. Deze stimulatie heeft nog een ander effect – CheB is inactief zonder fosforylering (het kan niet functioneren als methylesterase) en dit stelt het CheR-eiwit (een continu actief methyltransferase) in staat om te concurreren en vrijelijk de methyl-accepterende regio van het dimeer MCP (HAMP-domein) te methyleren. Deze methylering van het MCP heeft tot gevolg dat de autofosforyleringsgraad van het histidineresidu van CheA toeneemt. Daarom heeft het signaaltransductiesysteem controle over de flagellaire omkeerfrequentie in aanwezigheid van verhoogde of verlaagde stimuli, wat leidt tot een reeks “ren-en-terugkeer”-zwemen.

Tabel 2. Genen van MR-1 beschreven in de tekst.
Het is goed gedocumenteerd dat de accumulatie van MR-1 cellen in reactie op oplosbare elektronenacceptoren een vorm van energietaxis is, die afhangt van H+ flux en het tot stand brengen van een proton-motorkracht (Baraquet et al., 2009) (figuur 1). Een deel van deze respons omvat sneller zwemmen, zoals ook wordt gezien bij elektronenshuttles zoals riboflavine of antrachinon 2,6-disulfonaat (AQDS) (Bencharit en Ward, 2005; Harris et al., 2010; Li et al., 2012). In tegenstelling, de ophoping van cellen rond IEA, die is genoemd congregatie (Nealson et al., 1995), omvat zowel verhoogde zwemsnelheid bij contact met de IEA (de zogenaamde elektrokinese), en verhoogde zwemmen omkeringen bij een daling van de PMF. Zwemmen omkeringen mogelijk te maken meerdere voorbijgaande cel-IEA ontmoetingen (gedurende 1-100 ms), en de snelheid waarmee zwemmen cellen overgang naar onomkeerbare gehechtheid aan IEA tijdens de gemeente is nog niet gekwantificeerd.
Het mechanisme (s) die cellen gebruiken om IEA’s te lokaliseren blijven onduidelijk (Nealson et al., 1995; Bencharit en Ward, 2005; Harris et al., 2010). Vroege rapporten stelden dat het andere bestudeerde modelorganisme Geobacter zich rond IEA’s verzamelt door een gradiënt van gereduceerde metaalionen te detecteren (Childers et al., 2002), maar gereduceerde metaalionen zijn niet betrokken bij het detectiemechanisme dat door MR-1 wordt gebruikt (Bencharit and Ward, 2005). Bacteriële congregatie als reactie op gepoëleerde elektroden werd onlangs beschreven, wat wijst op de redoxdetectie, eerder dan de detectie van metaalionen, als de trigger voor de reactie (Harris et al., 2010). De positieve toegepaste potentialen op de elektrode (200-600 mV t.o.v. Ag/AgCl) veroorzaakten een samenkomst van MR-1 die vergelijkbaar was met die welke werd waargenomen bij metaaloxiden (Harris et al., 2012). Redoxpotentialen van MnO2-bevattende mineralen liggen tussen 400 en 600 mV vs. Ag/AgCl en aanvaarden gemakkelijker elektronen dan Fe(OH)3 mineralen, die een equivalent gepotentiaal van 100 tot 300 mV vs. Ag/AgCl bezitten (Burdige, 1993). In een eerdere studie werd verondersteld dat het karakteristieke zwemmen van MR-1 rond IEA wordt gereguleerd door twee zelf-sensorische chemotaxisreceptoren, methyl-accepterende proteïnen (MCP’s) met, Ca+-kanaal chemotaxisreceptor (Cache)-domein (SO_2240) en Per/Arnt/Sim (PAS)-domeinen (SO_1385), en door het chemotaxis signaaltransductie proteïne kinase CheA-3 (SO_3207) om de cel in staat te stellen het redoxpotentiaal, of elektronen accepterend vermogen van een oppervlak te monsteren (tabel 2 en figuur 1B) (Harris et al., 2012). Vóór deze studie waren de genen die verantwoordelijk zijn voor de motiliteit gedreven aanhechting van cellen op en rond IEA onbekend.
In dit rapport bestuderen we het congregatiemechanisme van verschillende Shewanella-stammen door het volgen van zwempatronen en celaanhechtingen aan MnO2 en Fe(OH)3. Naast het screenen van de WT en verschillende deletiemutanten van MR-1, werden ook andere Shewanella soorten (zie tabel 1) gescreend. Veel stammen bleken in staat te zijn tot congregatie rond zowel MnO2 als FeOH3, terwijl andere selectief reageerden op MnO2 (SB2B) of Fe(OH)3 (W3-18-1). Wanneer celhechting aan de minerale oppervlakken werd gecontroleerd, zagen we een sterke correlatie tussen het vermogen van de cel om samen te komen, en de gehechtheid van de cellen aan het IEA oppervlak. Vervolgens vergeleken we de genomen van deze soorten om kandidaat-genen te vinden die betrokken zijn bij het congregatiezwemmen, de accumulatie en het fenotype van celhechting in reactie op specifieke IEA-oppervlakken.
Results
Cell Accumulation and Attachment to Mineral Requires Chemotaxis and Extracellular Electron Transport Genes in S. oneidensis MR-1
De meeste van de vroege studies van energietaxis in MR-1 maakten gebruik van een methode van zwermplaat-tests (Nealson et al, 1995; Baraquet et al., 2009; Li et al., 2012). Hier bieden wij een meer diepgaande karakterisering van deze nog onbekende energie taxis mechanismen door gebruik te maken van een kwantitatieve methode van het volgen van het zwemmen van individuele cellen door vloeibare media in reactie op IEA en celaccumulatie na foto-bleken (CAAP) om celhechtingssnelheid te meten (Li et al., 2010). CAAP maakt gebruik van bestraling van een confocale microscoop UV-laser irreversibel verduisteren (doven van de fluorescentie van) GFP gelabelde cellen op en rond een bepaalde IEA. Door het kwantificeren van fluorescerende cellen als ze bewegen in de verduisterde zone, kan de tarieven van accumulatie en gehechtheid worden gekwantificeerd, en verschillende stammen vergeleken. Om te worden beschouwd als aangesloten, wordt de positie van de beweeglijke cel geverifieerd door het analyseren van een 3D-beeld van het gezichtsveld. Als de cel tot rust komt op het mineraaloppervlak, wordt de cel als “aangehecht” beschouwd. De sterkte van de bindingen tussen stationaire MR-1 cellen en oppervlakken werd gemeten met een optisch pincet in een afzonderlijke studie (Gross en El-Naggar, 2015). Deze methode maakt onderscheid tussen gerichte cel-attachment, en willekeurige elektrostatische gehechtheid.
Zoals blijkt uit figuur 2, de WT MR-1 cellen gehecht aan minerale oppervlak na 30 min met enkele grote afwijking in aantal, 2.655 ± 1.352 cellen / mm2. Gedurende deze tijd, de cellen vertonen “touch and go” zwemmen, het maken van voorbijgaande contact met de MnO 2 minerale oppervlak. Na 2 uur, het aantal nieuwe cellen aan het minerale oppervlak steeg tot 4.300 ± 584 cellen / mm2 van minerale oppervlak, terwijl extra beweeglijke cellen bleven samenkomen (Video’s S1, S2, en S17). Mutanten met drievoudige deleties in belangrijke EET genen mtrBC/omcA (SO_1776, SO_1778, SO_1779), enkelvoudige deletie van cymA (SO_4591) en de belangrijkste energie taxis chemoreceptor MCP Cache (SO_2240) zijn eerder in verband gebracht met zwemmen congregatie fenotype (Harris et al., 2012). Deletie mutanten (ΔcymA, Δmcp_cache, en ΔmtrBC/omcA) waren alle beweeglijk en in staat van het omkeren van zwemrichting, maar waren niet in staat van de gemeente en toonde weinig of geen gehechtheid aan MnO2 tijdens het experiment (figuur 2C en figuren S1A,B). WT accumulatie in gebleekte zone op t = 2 uur gemiddeld 4.300 ± 584 cellen / mm 2, terwijl Δmcp_cache, en ΔmtrBC/omcA mutanten accumuleren in verwaarloosbare aantallen (figuur 2C en S1CD).
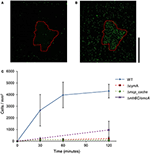
Figuur 2. Zwemmen MR-1 kan migreren in de richting van onoplosbare elektronenacceptor mineralen en hechten. Representatieve confocale fluorescentiemicroscopie beeld van WT MR-1 cellen op t = 0 en t = 120 min (A,B). Zwemmen WT MR-1 GFP-cellen werden geïntroduceerd om MnO 2 deeltje (rode stippellijn) in anaërobe afgesloten capillair. Op t = 0 alle cellen werden onomkeerbaar foto-gebleekt in een 250 bij 250 urn gebied rond deeltje (A). Fluorescerende cellen van buiten gebleekte zone die zwemmen in het frame en hechten aan minerale oppervlak werden vervolgens geteld. De zwarte verticale schaal balk aan de rechterkant vertegenwoordigt 100 um. Grafiek (C) vergelijkt WT MR-1 gehechtheid aan MnO2 deeltje meer dan 120 min met chemotaxis en extracellulaire elektron transfer schrapping mutanten (MR-1 Δmcp_cache, ΔcymA, en ΔmtrBC/omcA). De foutbalkjes omvatten 2 std deviations.
Characterizing Congregation rond IEA in andere Shewanella stammen
Negen stammen van Shewanella werden getest op hun vermogen om samen te komen rond minerale oppervlakken van MnO2 en Fe (OH) 3 met tijdreeksen assay van cel gehechtheid aan minerale (tabel 3 en Video’s S3-S14). Zoals bij MR-1 hadden alle negen onderzochte stammen een enkel polair flagellum en werd de zwemrichting omgekeerd door de rotatie van de flagellen om te keren (gegevens niet weergegeven). De zwemsporen binnen hetzelfde experiment werden gesorteerd in twee afzonderlijke groepen op basis van zwempad (tabel 3): degenen die contact onoplosbaar metaaloxide oppervlak (zwom binnen 2 pm) in vergelijking met degenen die geen contact (zwom > 2 pm). Contactmakende zwemmers die een significante toename in omkeerfrequentie en zwemsnelheid vertoonden dan de niet-contactmakende groep (P < 0,05) worden geclassificeerd als positief voor congregatiegedrag zoals aangeduid met superscriptletters (tabel 3).

Tabel 3. Bacteriën zwemsnelheid en omkeringsfrequentie rond metaaloxide mineralen.
Strains ANA3, CN32, en PV-4, in aanvulling op MR-1, waren positief voor gemeente rond zowel MnO2 en Fe(OH)3 (tabel 3, figuur 3; Video’s S1-S8). Zij vertoonden maximale omkeringspercentages wanneer zij zich tussen 5 en 40 μm van het MnO2- of FeOH3-deeltjesoppervlak bevonden (figuur 3). Drie stammen: OS217, NCIMB400, en OS155 waren niet beweeglijk op tijdstip t = 30 min-in reactie op Fe(OH)3 of MnO2 mineralen (tabel 3). Zo bedroeg de omkeerfrequentie van de zwemmende MR-1-cellen die in contact kwamen met MnO2 0,94 ± 0,53 omkeringen/s, terwijl de omkeerfrequentie van de niet-contacterende cellen 0,62 ± 0,73 omkeringen/s bedroeg. De snelheid nam ook toe in de contact makende groep MR-1-cellen, van 19,26 ± 11,2 μm/s in de contact makende groep tot 24,37 ± 6 μm/s in de contact makende groep. Omdat er zowel een statistisch significante toename van de zwemsnelheid en omkeringen zodat de cellen te blijven in de buurt van het metaaloxidedeeltje deze stam werd gezegd dat congregatie positief.
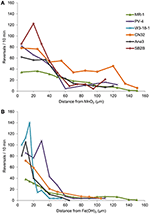
Figuur 3. (A,B) Omkeerfrequentie (y-as) van zwemmende Shewanella-cellen vs. afstand (x-as) van IEA-deeltje. De gemiddelde omkeerfrequenties van individuele gevolgd zwemmen cellen zijn verdeeld in bins van 5 pm langs de x-as voor experimenten met MnO2 (A) of Fe (OH)3 deeltje (B).
Shewanella spp. zwemmen tracks (30 s) die een voorkeur voor metaaloxide mineralen aangetoond zijn gemarkeerd in Figuren 4A-F. Figuur SB2B cellen vertoonden geen zwemmen reactie op Fe (OH) 3 (figuur 4D), terwijl het vertonen van actieve gemeente rond MnO2 deeltjes (figuur 4A). In tegenstelling, W3-18-1 cellen samengekomen rond Fe (OH) 3 deeltjes (figuur 4C), maar vertoonde een aanzienlijk verminderde zwemmen en omkering activiteit rond MnO2 (figuur 4F; Video’s S9, S14).
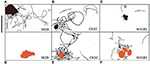
Figuur 4. (A-F) Paneel van drie Shewanella spp. zwemsporen (30 s) toonden een voorkeur aan voor metaaloxidemineralen. Het zwemgedrag van drie beweeglijke Shewanella spp. in reactie op MnO2 (bovenste rij-zwart gekleurd deeltje) en Fe(OH)3 (onderste rij-oranje gekleurd deeltje) werd bestudeerd. Congregatie gebeurt in anaërobe omstandigheden, zwemsporen zijn in zwart aangegeven. SB2B (links), CN32 (midden), en W3181 (rechts). Het zwemmen van de cellen werd met de hand gevolgd. De schaal balk op de linkerbovenhoek = 50 pm. De bruine kleur vorm vertegenwoordigt MnO2 en de oranje vertegenwoordigt Fe (OH) 3. Wanneer er geen beweeglijke cellen zijn gedetecteerd, wordt dit aangegeven door de afwezigheid van zwarte lijnen.
Discussie
Leden van het geslacht Shewanella bestaan uit heterotrofe, facultatieve aëroben die in staat zijn een breed scala van organische en anorganische stoffen als energiebron te benutten. Vele Shewanella-stammen zijn geïsoleerd uit de waterkolom en sedimenten op locaties over de hele wereld. Naast oplosbare elektronenacceptoren kunnen veel van deze organismen onder anoxische omstandigheden een breed assortiment van natuurlijk voorkomende onoplosbare metaaloxiden responderen.
In onze studie van congregatie gedrag in acht Shewanella soorten, gebruikten we cel tracking met computer analyse en tijdreeks assay van celhechting aan MnO2 of Fe(OH)3 om aan te tonen dat vijf van de acht shewanellae in staat waren dit gedrag CN32, ANA-3, W3-18-1, SB2B, en PV-4) en dat sommige stammen (W3-18-1, SB2B) vertonen een voorkeur voor een metaaloxide boven de andere onder deze omstandigheden (figuur 4). Dergelijke bevindingen zijn consistent met het idee dat deze bacteriën zich hebben aangepast aan de overheersende onoplosbare elektronenacceptor die wordt aangetroffen in de habitat waaruit zij zijn geïsoleerd. Bovendien kwamen drie Shewanella-stammen die uit de waterkolom werden geïsoleerd, niet samen als reactie op IEA. Of deze verschillen het gevolg zijn van de afwezigheid van genen die betrokken zijn bij EET (zoals bij de OS217) of van andere oorzaken, is een punt voor toekomstige studies.
In andere studies is bij nachtelijke groei van de Shewanella-stammen, waarbij de reductiesnelheden van verschillende metaaloxiden in een vergelijking van kop tot kop werden vergeleken, aangetoond dat zij bij voorkeur metaaloxide uit hun milieu-niche reduceren (Bretschger et al., 2007). De hier gepresenteerde resultaten zijn consistent met de hypothese dat Shewanella-soorten een samenlevings- en aanhechtingsgedrag hebben geëvolueerd dat consistent is met de milieus waaruit zij werden geïsoleerd (Tabel 3). Zo lijkt W3-18-1 een significante neiging tot congregatie te vertonen in aanwezigheid van en hechting aan Fe(OH)3 mineralen die inheems zijn in het continentaal plat van de Stille Oceaan (Harris et al., 2012), terwijl SB2B de voorkeur geeft aan MnO2 (Venkateswaran et al., 1998), vergelijkbaar met dat van het sediment van de Amazonerivier.
De genomische vergelijking in tabel 1 suggereert dat de aanwezigheid van octaheme cytochroom (SO_4142) belangrijk kan zijn voor het zwemmen in reactie op relatief “lage” redoxpotentialen (100-300 mV vs. Ag/AgCl) van Fe(OH)3. Terwijl mtrF (SO_1780) nodig kan zijn voor de respons op relatief “hogere” redoxpotentialen van MnO2 (tussen 400 en 600 mV t.o.v. Ag/AgCl). Deze hypothese kan vervolgens worden getest met deletiemutanten in MR-1. Dit werk vormt een aanvulling op eerdere werk van Harris et al. (2012), door te onthullen dat genen die betrokken zijn bij het netto-effect van de motiliteit gedrag in de richting van IEA mineralen of elektroden is cumulatief door middel van relevante tijdschalen van 1-30 min (figuur 2) (Harris et al., 2012). Onze bevindingen met CAAP confocale microscopie techniek, toonde aan dat de gemeente leidt tot een langzame migratie van cellen in de richting van IEA mineralen met uiteindelijke gehechtheid. Zo wordt dit gedrag toepasselijk congregatie genoemd, omdat het de verzameling of accumulatie van beweeglijke bacteriën rond IEA beschrijft. We toonden hier aan dat de genen mcp cache en mtrBC/omcA essentieel zijn voor het accumulatie- en aanhechtingsfenotype in MR-1, naast het feit dat ze verantwoordelijk zijn voor de karakteristieke zwempatronen van verhoogde snelheid en ren-en-terug-type gedrag dat eerder werd geïdentificeerd (Harris et al, 2012).
De resultaten van de experimenten met ΔcymA, ΔmtrBC/omcA, en Δmcp_cache mutanten in MR-1 tonen een remming van accumulatie, aanhechting en samenlevingsgedrag in reactie op alle IEA’s. Daarom kan worden verondersteld dat de aanwezigheid van homologe EET genen (cymA, mtrB, mtrC, omcA), en methyl accepterend chemotaxis gen (mcp_cache) de fenotypische responsen bepaalt die we zien bij andere Shewanella spp. (Tabel 3). Genetische vergelijking van stammen, die kunnen reageren op “lagere” redoxpotentiaal IEA, geeft aan dat perifere buitenmembraan octaheem cytochromen (zoals SO_4142) een rol kunnen spelen in het reageren op “lagere” redoxpotentiaal IEA. Vandaar dat deze waarnemingen over aanhoudend zwemmen rond specifieke mineralen overeenkwamen met specifieke genotypen van de verschillende soorten.
Tijdens veel MFC- (Kotloski en Gralnick, 2013) en metaalreducerende batchcultuurexperimenten werd extracellulaire elektronentransfer-gemedieerde energietaxis, of congregatievermogen, niet gemeten (Kotloski en Gralnick, 2013). In metaaloxidereductieproeven kunnen de 3D-distributie van cellen en de beweeglijkheid van cellen de metaalreductie sterk beïnvloeden als gevolg van de snelheid van het schudapparaat, de afmetingen van de kweekfles of het mengen. Congregatie gedrag direct van invloed cel gehechtheid aan IEAs. De relatie tussen congregatie en nanodraad-achtige aanhangselvorming is nog onbekend (Pirbadian et al., 2015), hoewel de motiliteit en congregatie parameters zijn zelden gecontroleerd in deze studies (Gorby et al., 2005; Reguera et al., 2005). Het transformeren van deze kerngemeentegenen (cymA, mtrB, mtrA, omcA, mcp_pas, en mcp_cache) in andere bacteriesoorten, met enkelpolig flagellum, kan op een dag de bioremediëringsmogelijkheden verbeteren door bacteriële aanhechting en kolonisatie te kunnen induceren van oppervlakken die anders moeilijk of onmogelijk zouden zijn.
Materialen en methoden
Kweek en stammen
MR-1 en verschillende deletiemutanten afkomstig van MR-1 werden in deze studie onderzocht (tabel 2) (Beliaev en Saffarini, 1998; Myers en Myers, 2002). De stammen werden vanuit glycerolvoorraden die bij -80°C waren bewaard, geënt op Luria-Bertani-platen (LB) en ’s nachts bij 30°C gekweekt. Individuele kolonies werden vervolgens geselecteerd en geïnoculeerd in 5 ml van gedefinieerde minimale media (M1) (Bretschger et al., 2007), aangevuld met 18 mM lactaat als energiebron (Bretschger et al., 2007) in buisjes van 15 ml (VWR International LLC, Randor, Pennsylvania, USA) en horizontaal geïncubeerd in een shaker (180 rpm) gedurende 48 uur bij 30 °C. De optische dichtheid werd gemeten met een spectrofotometer (Unico 1100RS spectrofotometer, Dayton, New Jersey, USA). Cellen werden bemonsterd bij een OD600 van 0,5 ± 0,2 (na ~48 h). Bij zwemexperimenten werden vijf milliliter culturen bemonsterd wanneer de cellen een OD van 0,4 hadden bereikt, gemengd met mangaan- of ijzeroxiden, en ingebracht in een glazen capillair (0,02 × 0,20 mm) (Vitrocom, Mountain Lakes, New Jersey, VS) dat vervolgens werd verzegeld met vacuümvet zoals eerder beschreven (Harris et al, 2010).
Minerale Synthese
De Fe (OH) 3 voorraadoplossing werd bereid volgens het protocol door Cornell en Schwertmann en vervolgens geverifieerd door X-ray defractie (Schwertmann en Cornell, 2008). Deze bereiding van colloïdaal MnO2 begon met 8 g KMnO4 opgelost in 200 mL, met gebruikmaking van alle mogelijke voorzorgsmaatregelen. De oplossing werd voortdurend gemengd met behulp van een magnetische roerstaaf op hoog en verwarmd tot onder de kooktemperatuur. Vervolgens werd 5 mL 10 M natriumhydroxide toegevoegd om het bij de reactie ontstane zuur te neutraliseren. In een aparte kolf werd 15 g mangaanchloride opgelost in 75 mL gedestilleerd water. De oplossing werd vervolgens langzaam gemengd met de permanganaatoplossing (in een chemische zuurkast) gedurende 75 minuten. Na afkoeling werd de oplossing vervolgens gewassen door centrifugeren en vijfmaal gespoeld met gedeïoniseerd water (DI) (18 Meg-Ohm cm). Het uiteindelijke neerslag werd met behulp van een vacuümfilter in een schone bank gedroogd en gedurende 36 uur gedroogd. De resulterende mineralen werden via röntgendiffractie geanalyseerd om de productie van Fe(OH)3 en MnO2 te bevestigen (Bretschger et al., 2007; Salas et al., 2010).
Gesuspendeerde minerale deeltjes werden met cultuur gemengd in een eindconcentratie van 300 mg/ml MnO2 of Fe(OH)3. De cellen werden vervolgens capillair ondergedompeld in rechthoekige capillaire buisjes (0,02 × 0,20 mm) (Vitrocom, Mountain Lakes, New Jersey, USA). Buizen werden verzegeld met silicium vacuüm vet (Dow Corning, Midland, Milwaukie, USA) en waargenomen met lichtmicroscopie, fluorescentie microscopie, en confocale microscopie.
Cell Accumulation after Photo-Bleaching (CAAP) Time-Lapse Experiments
Alle fluorescent gelabelde stammen (GFP) werden getransformeerd zoals eerder beschreven (McLean et al, 2008) en vervolgens aëroob gekweekt op een gedefinieerd minimaal medium met 25 ug / ml kanamycine en 18 mM lactaat gedurende 48 uur bij 30 ° C. Vijf milliliter culturen werden bemonsterd wanneer de cellen een OD van 0,4 hadden bereikt, gemengd met mangaan- of ijzeroxiden, en ingebracht in een glazen capillair (0,02 × 0,20 mm) (Vitrocom, Mountain Lakes, New Jersey, USA) dat vervolgens werd verzegeld met vacuümvet zoals eerder beschreven (Harris et al., 2010). GFP gelabeld WT MR-1, ΔcymA, Δmcp_cache, en ΔmtrBC/omcA cellen werden gebleekt met behulp van maximale lichtintensiteit instellingen met 60X lenzen, van een omgekeerde Leica TCS SPE confocale microscoop (Wetzlar, Duitsland) gedurende 15 minuten. Om ervoor te zorgen dat bleken plaatsgevonden, time-lapse vastgelegd een scherm gebied (250 × 250 urn geselecteerd gebied) elke minuut tot de oorspronkelijke cellen donker en de omliggende cellen bleef helder fluorescerende. Beelden werden vervolgens vastgelegd met 588 nm excitatie en 530 nm emissie. Een time-lapse video van het gehele deel van de buis werd vastgelegd met behulp van Leica Imaging software en de “auto focus”-functie voor de komende 3 uur. Cellen werden ook waargenomen onder doorvallend licht-modus om te controleren of gebleekte cellen beweeglijk en intact waren. Een aparte negatieve controle, met GFP gelabeld Δmcp_cache, werd vastgelegd voor 3 h. Geen cellen werden gezien accumuleren in de donkere zone in deze negatieve controle, noch gebleekte cellen herstellen GFP fluorescentie. De reactie van het gehele capillair (hoogte van 30 pm) werd vastgelegd met time-lapse fotografie en de som van de cellen in alle 20 z-as stacks werd bepaald (met behulp van computer analyse methode hieronder) voor elk tijdstip.
Microscopie Capture van celbewegingen
De methoden voor bacteriële tracking en analyse waren identiek aan eerdere studies gebruikt voor figuur 3 en opgenomen in tabel 3 (Harris et al., 2012). Zowel computer en handmatige bacteriële zwemmen tracks werden gestandaardiseerd met behulp van een schaal liniaal (100 pm). Van elk experiment, de totale zwemactiviteit binnen het videoframe-equivalent aan een 107 × 193 urn gezichtsveld werd opgenomen en de video werd tijd genormaliseerd om zwemmen snelheden in um / s te geven. Verschillende parameters werden gemeten voor elke bacteriële zwemmen track, zoals de totale afstand verplaatst, de tijd van het spoor sinds de bacteriën voor het eerst verschijnen en verdwijnen, het aantal omkeringen, de afstand tussen elke omkering en het metaaloxide, en de afstand tussen het metaaloxide en het begin van de bacteriën track.
Zwemmen Analysis
Cellen werden bijgehouden met de hand van video-gegevens (30 frames / s), frame-voor-frame, en gemeten door ImageJ beeldanalyse software. Data-ingangen zijn de zwemsnelheid, de uitgangspositie van de bacteriën ten opzichte van de dichtstbijzijnde IEA oppervlak en de positie van elke bacteriële omkering gebeurtenis werd geïdentificeerd en gelogd met betrekking tot de afstand van de dichtstbijzijnde IEA oppervlak (figuur 3 en tabel 3). Voor het bepalen van de zwemactiviteit na contact met IEA, werden de zwemmende cellen verdeeld in twee groepen voor analyse: cellen die zwommen binnen 2 pm van een deeltje werden beschouwd als “contact” en die niet zwemmen binnen 2 pm van het deeltje oppervlak werden beschouwd als “niet-contacterende”. In aanvulling op de hand tracking methoden hierboven beschreven, werd experimentele data vervolgens computer gesorteerd en geanalyseerd door een algoritme eerder beschreven om de berekende zwemmen gegevens (Crocker en Grier, 1996; Harris et al., 2012; Harris, 2013) opleveren. Om Figuren 3, 4 te produceren, werden alle experimentele gegevens van vier afzonderlijke biologische replicaten gecombineerd, gepoold en gemiddeld in bins op basis van afstand van de IEA deeltje. Omdat deze gegevens werden gepoold in afstanden, zijn foutbalken overlappende; geen vergelijking van omkering frequentie, op een bepaalde afstand kan worden gemaakt tussen stammen, zoals deze methode van visualisatie sluit experiment identifiers.
Author Contributions
HH: ontworpen en uitgevoerd de experimenten; HH en IS-A: schreef en herzien paper; ES, ME-N, en JM: hielp bij het ontwerpen experimenten; WT: hielp bij het uitvoeren van de data-analyse; JM en KN: herzien het manuscript.
Conflict of Interest Statement
De auteurs verklaren dat het onderzoek werd uitgevoerd in de afwezigheid van commerciële of financiële relaties die kunnen worden opgevat als een potentieel belangenconflict.
Acknowledgments
Speciale dank aan Mandy J. Ward en Gijs Kuenen voor advies over onderzoek en experiment design. Wij danken Meaghan Sullivan voor haar handmatige tracking analyses. Wij danken Cecile Jourlin-Castelli, Samantha Reed, Jun Li, en David Culley voor het leveren van de Δmcp_cache, ΔmtrBC/omcA, en ΔcymA mutanten. Financiering voor dit werk werd verstrekt door een Air Force Office of Scientific Research Award . IS-A onderzoek werd ondersteund door de NASA Planetaire Biologie Stage fellowship.
Aanvullend materiaal
Het aanvullend materiaal voor dit artikel kan online worden gevonden op: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02568/full#supplementary-material
Alexandre, G., Greer-Phillips, S., and Zhulin, I. B. (2004). Ecologische rol van energietaxi’s in micro-organismen. FEMS Microbiol. Rev. 28, 113-126. doi: 10.1016/j.femsre.2003.10.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Baraquet, C., Théraulaz, L., Iobbi-Nivol, C., Méjean, V., and Jourlin-Castelli, C. (2009). Onverwachte chemoreceptoren mediëren energietaxis naar elektronenacceptoren in Shewanella oneidensis. Mol. Microbiol. 73, 278-290. doi: 10.1111/j.1365-2958.2009.06770.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Beliaev, A. S., and Saffarini, D. A. (1998). Shewanella putrefaciens mtrB codeert voor een buitenmembraaneiwit dat nodig is voor Fe(III) en Mn(IV) reductie. J. Bacteriol. 180, 6292-6297.
PubMed Abstract | Google Scholar
Bencharit, S., and Ward, M. J. (2005). Chemotactische reacties op metalen en anaerobe elektronenacceptoren in Shewanella oneidensis MR-1. J. Bacteriol. 187, 5049-5053. doi: 10.1128/JB.187.14.5049-5053.2005
PubMed Abstract | CrossRef Full Text | Google Scholar
Bond, D. R., and Lovley, D. R. (2003). Elektriciteitsproductie door Geobacter sulfurreducens bevestigd aan elektroden. Application. Environ. Microbiol. 69, 1548-1555. doi: 10.1128/AEM.69.3.1548-1555.2003
PubMed Abstract | CrossRef Full Text | Google Scholar
Bowman, J. P., McCammon, S. A., Nichols, D. S., Skerratt, J. H., Rea, S. M., Nichols, P. D., et al. (1997). Shewanella gelidimarina sp. nov. and Shewanella frigidimarina sp. nov., novel antarctic species with the ability to produce eicosapentaenoic acid (20:5ω3) and grow anaerobically by dissimilatory Fe(III) reduction. Int. J. Syst. Bacteriol. 47, 1040-1047. doi: 10.1099/00207713-47-4-1040
PubMed Abstract | CrossRef Full Text | Google Scholar
Bretschger, O. (2008). Electron Transfer Capability and Metabolic Processes of the Genus Shewanella with Applications to the Optimization of Microbial Fuel Cells. University of Southern California, ProQuest Dissertations Publishing, 3325185.
Google Scholar
Bretschger, O., Obraztsova, A., Sturm, C. A., Chang, I. S., Gorby, Y. A., Reed, S. B., et al. (2007). Current production and metal oxide reduction by Shewanella oneidensis MR-1 wild type and mutants. Application. Environ. Microbiol. 73:7003. doi: 10.1128/AEM.01087-07
PubMed Abstract | CrossRef Full Text | Google Scholar
Brettar, I., Christen, R., and Höfle, M. G. (2002). Shewanella denitrificans sp nov., a vigorously denitrifying bacterium isolated from the oxic-anoxic interface of the Gotland Deep in the central Baltic Sea. Int. J. Syst. Evol. Microbiol. 52, 2211-2217. doi: 10.1099/00207713-52-6-2211
PubMed Abstract | CrossRef Full Text | Google Scholar
Burdige, D. J. (1993). The biogeochemistry of manganese and iron reduction in marine sediments. Earth Sci. Rev. 35, 249-284. doi: 10.1016/0012-8252(93)90040-E
CrossRef Full Text | Google Scholar
Champine, J. E., Underhill, B., Johnston, J. M., Lilly, W. W., and Goodwin, S. (2000). Electron transfer in the dissimilatory iron-reducing bacterium Geobacter metallireducens. Anaerobe 6, 187-196. doi: 10.1006/anae.2000.0333
CrossRef Full Text | Google Scholar
Childers, S. E., Ciufo, S., and Lovley, D. R. (2002). Geobacter metallireducens krijgt toegang tot onoplosbaar Fe(III)oxide door chemotaxis. Nature 416, 767-769. doi: 10.1038/416767a
PubMed Abstract | CrossRef Full Text | Google Scholar
Coursolle, D., and Gralnick, J. A. (2010). Modulariteit van de Mtr respiratoire pathway van Shewanella oneidensis stam MR-1. Mol. Microbiol. 77, 995-1008. doi: 10.1111/j.1365-2958.2010.07266.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Crocker, J. C., and Grier, D. G. (1996). Methods of digital video microscopy for colloidal studies. J. Colloid Interface Sci. 179, 298-310. doi: 10.1006/jcis.1996.0217
CrossRef Full Text | Google Scholar
El-Naggar, M. Y., Wanger, G., Leung, K. M., Yuzvinsky, T. D., Southam, G., Yang, J., et al. (2010). Elektrisch transport langs bacteriële nanodraden van Shewanella oneidensis MR-1. Proc. Natl Acad. Sci. U.S.A. 107, 18127-18131. doi: 10.1073/pnas.1004880107
PubMed Abstract | CrossRef Full Text | Google Scholar
Fredrickson, J. K., Romine, M. F., Beliaev, A. S., Auchtung, J. M., Driscoll, M. E., Gardner, T. S., et al. (2008). Naar een milieusysteembiologie van Shewanella. Nat. Rev. Microbiol. 6, 592-603. doi: 10.1038/nrmicro1947
PubMed Abstract | CrossRef Full Text | Google Scholar
Fredrickson, J. K., Zachara, J. M., Kennedy, D. W., Dong, H., Onstott, T. C., Hinman, N. W., et al. (1998). Biogene ijzermineralisatie bij de dissimilatoire reductie van waterhoudend ijzeroxide door een grondwaterbacterie. Geochim. Cosmochim. Acta 62, 3239-3257. doi: 10.1016/S0016-7037(98)00243-9
CrossRef Full Text | Google Scholar
Gao, H., Obraztova, A., Stewart, N., Popa, R., Fredrickson, J. K., Tiedje, J. M., et al. (2006). Shewanella loihica sp. nov., geïsoleerd uit ijzerrijke microbiële matten in de Stille Oceaan. Int. J. Syst. Evol. Microbiol. 56, 1911-1916. doi: 10.1099/ijs.0.64354-0
PubMed Abstract | CrossRef Full Text | Google Scholar
Gorby, Y. A., Beveridge, T. J., and Wiley, W. R. (2005). “Composition, reactivity, and regulation of extracellular metal-reducing structures (nanowires) produced by dissimilatory metal reducing bacteria,” in Annual NABIR PI Meeting (Warrenton, VA).
Google Scholar
Gorby, Y. A., Yanina, S., McLean, J. S., Rosso, K. M., Moyles, D., Dohnalkova, A., et al. (2006). Elektrisch geleidende bacteriële nanodraden geproduceerd door Shewanella oneidensis stam MR-1 en andere micro-organismen. Proc. Natl. Acad. Sci. U.S.A. 103, 11358-11363. doi: 10.1073/pnas.0604517103
PubMed Abstract | CrossRef Full Text | Google Scholar
Gross, B. J., and El-Naggar, M. Y. (2015). Een gecombineerde elektrochemische en optische trapping platform voor het meten van enkele cel respiratie snelheden op elektrode interfaces. Rev. Sci. Instrum. 86:064301. doi: 10.1063/1.4922853
PubMed Abstract | CrossRef Full Text | Google Scholar
Harris, H. W. (2013). Identificatie van een nieuw Bacterieel Sensing Mechanisme: Karakterisering van Bacteriële Onoplosbare Elektronen Acceptor Sensing. University of Southern California.
Google Scholar
Harris, H. W., El-Naggar, M. Y., Bretschger, O., Ward, M. J., Romine, M. F., Obraztsova, A. Y., et al. (2010). Electrokinesis is een microbieel gedrag dat extracellulair elektronentransport vereist. Proc. Natl. Acad. Sci. U.S.A. 107, 326-331. doi: 10.1073/pnas.0907468107
PubMed Abstract | CrossRef Full Text | Google Scholar
Harris, H. W., Naggar, M. Y., and Nealson, K. H. (2012). Shewanella oneidensis MR-1 chemotaxis eiwitten en electron-transport chain componenten essentieel voor congregatie nabij onoplosbare electron acceptoren. Biochem. Soc. Trans. 40, 1167-1177. doi: 10.1042/BST20120232
PubMed Abstract | CrossRef Full Text | Google Scholar
Hau, H. H., and Gralnick, J. A. (2007). Ecologie en biotechnologie van het genus Shewanella. Annu. Rev. Microbiol. 61, 237-258. doi: 10.1146/annurev.micro.61.080706.093257
PubMed Abstract | CrossRef Full Text | Google Scholar
Hsu, L., Masuda, S. A., Nealson, K. H., and Pirbazari, M. (2012). Evaluatie van microbiële brandstofcel Shewanella biokathoden voor de behandeling van chromaatverontreiniging. R. Soc. Chem. Adv. 2, 5844-5855. doi: 10.1039/c2ra20478a
CrossRef Full Text | Google Scholar
Kan, J., Hsu, L., Cheung, A. C. M., Pirbazari, M., and Nealson, K. H. (2011). Stroomproductie door bacteriële gemeenschappen in microbiële brandstofcellen verrijkt uit afvalwaterslib met verschillende elektronendonoren. Environ. Sci. Technol. 45, 1139-1146. doi: 10.1021/es102645v
PubMed Abstract | CrossRef Full Text | Google Scholar
Kato, S., Nakamura, R., Kai, F., Watanabe, K., and Hashimoto, K. (2010). Respiratoire interacties van bodembacteriën met (half)geleidende ijzer-oxide mineralen. Environ. Microbiol. 12, 3114-3123. doi: 10.1111/j.1462-2920.2010.02284.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Kotloski, N. J., and Gralnick, J. A. (2013). Flavin electron shuttles dominate extracellular electron transfer by Shewanella oneidensis. MBio 4:e00553. doi: 10.1128/mBio.00553-12
PubMed Abstract | CrossRef Full Text | Google Scholar
Kracke, F., Vassilev, I., and Krömer, J. O. (2015). Microbieel elektronentransport en energiebehoud – de basis voor het optimaliseren van bio-elektrochemische systemen. Front. Microbiol. 6:575. doi: 10.3389/fmicb.2015.00575
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, J., Go, A. C., Ward, M. J., and Ottemann, K. M. (2010). De chemische-in-plug bacteriële chemotaxis assay is gevoelig voor vals-positieve reacties. BMC Res. Notes 3:77. doi: 10.1186/1756-0500-3-77
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, J., Romine, M. F., and Ward, M. J. (2007). Identification and analysis of a highly conserved chemotaxis gene cluster in Shewanella species. FEMS Microbiol. Lett. 273, 180-186. doi: 10.1111/j.1574-6968.2007.00810.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, R., Tiedje, J. M., Chiu, C., and Worden, R. M. (2012). Soluble electron shuttles can mediate energy taxis to insoluble electron acceptors. Environ. Sci. Technol. 46, 2813-2820. doi: 10.1021/es204302w
PubMed Abstract | CrossRef Full Text | Google Scholar
Lovley, D. R., Coates, J. D., Blunt-Harris, E. L., Phillips, E. J. P., and Woodward, J. C. (1996). Humusstoffen als elektronenacceptoren voor microbiële respiratie. Nature 382, 445-448. doi: 10.1038/382445a0
CrossRef Full Text | Google Scholar
Lovley, D. R., Giovannoni, S. J., White, D. C., Champine, J. E., Phillips, E. J., Gorby, Y. A., et al. (1993). Geobacter metallireducens gen. nov. sp. nov., a microorganism capable of coupling the complete oxhidation of organic compounds to the reduction of iron and other metals. Arch. Microbiol. 159, 336-344. doi: 10.1007/BF00290916
PubMed Abstract | CrossRef Full Text | Google Scholar
Lovley, D. R., Holmes, D. E., and Nevin, K. P. (2004). Dissimilatoire reductie van Fe(III) en Mn(IV). Adv. Microb. Physiol. 49, 219-286. doi: 10.1111/j.1462-2920.2004.00593.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Marsili, E., Baron, D. B., Shikhare, I. D., Coursolle, D., Gralnick, J. A., Bond, D. R., et al. (2008). Shewanella scheidt flavines af die extracellulaire elektronenoverdracht bewerkstelligen. Proc. Natl. Acad. Sci. U.S.A. 105, 3968-3973. doi: 10.1073/pnas.0710525105
PubMed Abstract | CrossRef Full Text | Google Scholar
McLean, J. S., Majors, P. D., Reardon, C. L., Bilskis, C. L., Reed, S. B., Romine, M. F., et al. (2008). Onderzoek naar structuur en metabolisme binnen Shewanella oneidensis MR-1 biofilms. J. Microbiol. Methods 74, 47-56. doi: 10.1016/j.mimet.2008.02.015
PubMed Abstract | CrossRef Full Text | Google Scholar
Meyer, T. E., Tsapin, A. I., Vandenberghe, I., de Smet, L., Frishman, D., Nealson, K. H., et al. (2004). Identification of 42 possible cytochrome C genes in the Shewanella oneidensis genome and characterization of six soluble cytochromes. Omics 8, 57-77. doi: 10.1089/153623104773547499
PubMed Abstract | CrossRef Full Text | Google Scholar
Mitchell, A. C., Peterson, L., Reardon, C. L., Reed, S. B., Culley, D. E., Romine, M. R., et al. (2012). Role of outer membrane c-type cytochromes MtrC and OmcA in Shewanella oneidensis MR-1 cell production, accumulation, and detachment during respiration on hematite. Geobiology 10, 355-370. doi: 10.1111/j.1472-4669.2012.00321.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Murray, A. E., Lies, D., Li, G., Nealson, K., Zhou, J., and Tiedje, J. M. (2001). DNA/DNA hybridisatie op microarrays onthult gen-specifieke verschillen tussen nauw verwante microbiële genomen. Proc. Natl. Acad. Sci. U.S.A. 98, 9853-9858. doi: 10.1073/pnas.171178898
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Myers, J. M. (1997). Cloning and sequence of cymA, a gene encoding a tetraheme cytochrome c required for reduction of iron(III), fumarate, and nitrate by Shewanella putrefaciens MR-1. J. Bacteriol. 179, 1143-1152. doi: 10.1128/jb.179.4.1143-1152.1997
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Myers, J. M. (2002). MtrB Is nodig voor de juiste incorporatie van de cytochromen OmcA en OmcB in het buitenmembraan van Shewanella putrefaciens MR-1. Appl. Environ. Microbiol. 68, 5585-5594. doi: 10.1128/AEM.68.11.5585-5594.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Nealson, K. H. (1988a). Bacterial manganese reduction and growth with manganese oxide as the sole electron acceptor. Science 240, 1319-1321. doi: 10.1126/science.240.4857.1319
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Nealson, K. H. (1988b). Microbiële reductie van mangaanoxiden: Interacties met ijzer en zwavel. Geochim. Cosmochim. Acta 52, 2727-2732. doi: 10.1016/0016-7037(88)90041-5
CrossRef Full Text | Google Scholar
Myers, J. M., and Myers, C. R. (2001). Role for outer membrane cytochromes OmcA and OmcB of Shewanella putrefaciens MR-1 in reduction of manganese dioxide. Appl. Environ. Microbiol. 67, 260-269. doi: 10.1128/AEM.67.1.260-269.2001
PubMed Abstract | CrossRef Full Text | Google Scholar
Nealson, K. H., Moser, D. P., and Saffarini, D. A. (1995). Anaerobic electron acceptor chemotaxis in Shewanella putrefaciens. Appl. Environ. Microbiol. 61, 1551-1554.
PubMed Abstract | Google Scholar
Okamoto, A., Kalathil, S., Deng, X., Hashimoto, K., Nakamura, R., and Nealson, K. H. (2014). Cell-secreted flavins bound to membrane cytochromes dictate electron transfer reactions to surfaces with divers charge and pH. Sci. Rep. 4:5628. doi: 10.1038/srep05628
PubMed Abstract | CrossRef Full Text | Google Scholar
Pirbadian, S., Barchinger, S. E., Leung, K. M., Byun, H. S., Jangir, Y., Bouhenni, R. A., et al. (2015). Bacteriële nanodraden van Shewanella oneidensis MR-1 zijn buitenmembraan en periplasmatische uitbreidingen van de extracellulaire elektronentransportcomponenten. Biophys. J. 108:368a. doi: 10.1016/j.bpj.2014.11.2016
CrossRef Full Text | Google Scholar
Reguera, G., McCarthy, K. D., Mehta, T., Nicoll, J. S., Tuominen, M. T., Lovely, D. R., et al. (2005). Extracellulaire elektronenoverdracht via microbiële nanodraden. Nature 435, 1098-1101. doi: 10.1038/nature03661
PubMed Abstract | CrossRef Full Text | Google Scholar
Salas, E. C., Berelson, W. M., Hammond, D. E., Kampf, A. R., and Nealson, K. H. (2010). De impact van bacteriestammen op de producten van dissimilatoire ijzerreductie. Geochim. Cosmochim. Acta 74, 574-583. doi: 10.1016/j.gca.2009.10.039
PubMed Abstract | CrossRef Full Text | Google Scholar
Saltikov, C. W., Cifuentes, A., Venkateswaran, K., and Newman, D. K. (2003). The ars detoxification system is advantageous but not required for As(V) respiration by the genetically tractable Shewanella species strain ANA-3. Appl. Environ. Microbiol. 69, 2800-2809. doi: 10.1128/AEM.69.5.2800-2809.2003
CrossRef Full Text | Google Scholar
Schwalb, C., Chapman, S. K., and Reid, G. A. (2003). The tetraheme cytochrome CymA is required for anaerobic respiration with dimethyl sulfoxide and nitrite in Shewanella oneidensis. Biochemistry 42, 9491-9497. doi: 10.1021/bi034456f
PubMed Abstract | CrossRef Full Text | Google Scholar
Schwertmann, U., and Cornell, R. M. (2008). IJzeroxiden in het Laboratorium. Weinheim: John Wiley & Sons.
Google Scholar
Shi, L., Richardson, D. J., Wang, Z., Kerisit, S. N., Rosso, K. M., Zachara, J. M., et al. (2009). De rol van cytochromen in het buitenmembraan van Shewanella en Geobacter in extracellulaire elektronenoverdracht. Environ. Microbiol. Rep. 1, 220-227. doi: 10.1111/j.1758-2229.2009.00035.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Venkateswaran, K., Dollhopf, M. E., Aller, R., Stackebrandt, E., and Nealson, K. H. (1998). Shewanella amazonensis sp. nov., a novel metal-reducing facultative anaerobe from Amazonian shelf muds. Int. J. Syst. Bacteriol. 48, 965-972.
PubMed Abstract | Google Scholar
Venkateswaran, K., Moser, D. P., Dollhopf, M. E., Lies, D. P., Saffarini, D. A., MacGregor, B. J., et al. (1999). Polyphasic taxonomy of the genus Shewanella and description of Shewanella oneidensis sp. nov. Int. J. Syst. Bacteriol. 49, 705-724. doi: 10.1099/00207713-49-2-705
PubMed Abstract | CrossRef Full Text | Google Scholar
Ziemke, F., Höfle, M. G., Lalucat, J., and Rossellö-Mora, R. (1998). Reclassification of Shewanella putrefaciens Owen’s genomic group II as Shewanella baltica sp. nov. Int. J. Syst. Bacteriol. 48, 179-186. doi: 10.1099/00207713-48-1-179
PubMed Abstract | CrossRef Full Text | Google Scholar