Jak pokrótce opisano w poprzednim rozdziale, neuroprzekaźniki są sygnałami chemicznymi uwalnianymi z presynaptycznych zakończeń nerwowych do szczeliny synaptycznej. Późniejsze wiązanie neuroprzekaźników ze specyficznymi receptorami na neuronach postsynaptycznych (lub innych klasach komórek docelowych) przejściowo zmienia właściwości elektryczne komórek docelowych, prowadząc do ogromnej różnorodności efektów postsynaptycznych (patrz Rozdziały 7 i 8).
Pogląd, że informacja elektryczna może być przekazywana z jednego neuronu do drugiego za pomocą sygnalizacji chemicznej był przedmiotem intensywnej debaty w pierwszej połowie XX wieku. Kluczowy eksperyment, który poparł tę ideę, został przeprowadzony w 1926 roku przez niemieckiego fizjologa Otto Loewi. Działając na podstawie pomysłu, który rzekomo przyszedł mu do głowy w środku nocy, Loewi udowodnił, że elektryczna stymulacja nerwu błędnego spowalnia bicie serca poprzez uwalnianie sygnału chemicznego. Wyizolował i perfundował serca dwóch żab, monitorując tempo ich bicia (rysunek 6.1). Istotą jego eksperymentu było zebranie płynu perfuzyjnego przepływającego przez stymulowane serce i przeniesienie go do drugiego serca. Mimo, że drugie serce nie było stymulowane, jego bicie również zwolniło, co świadczy o tym, że nerw błędny reguluje rytm serca poprzez uwalnianie substancji chemicznej, która gromadzi się w perfuzji. Pierwotnie określany jako „substancja błędnika”, czynnik ten został później uznany za acetylocholinę (ACh), która z biegiem lat stała się najdokładniej zbadanym neuroprzekaźnikiem. ACh działa nie tylko w sercu, ale w wielu postsynaptycznych celach w centralnym i obwodowym układzie nerwowym, przede wszystkim w złączu nerwowo-mięśniowym mięśni prążkowanych i w trzewnym układzie ruchowym (zob. rozdziały 5 i 21).
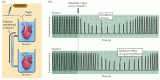
Rysunek 6.1
Doświadczenie Loewiego demonstrujące chemiczną neurotransmisję. (A) Schemat układu doświadczalnego. (B) Gdy nerw błędny izolowanego serca żaby był stymulowany, częstość akcji serca spadała (górny panel). Jeśli płyn perfuzyjny ze stymulowanego serca (więcej…)
Na przestrzeni lat pojawiło się wiele formalnych kryteriów, które definitywnie identyfikują daną substancję jako neuroprzekaźnik (Ramka A). Niemniej jednak, identyfikacja neuroprzekaźników aktywnych w każdej konkretnej synapsie pozostaje trudnym przedsięwzięciem, a w przypadku wielu synaps (szczególnie w mózgu), charakter neuroprzekaźnika nie jest dobrze ustalony. Substancje, które nie spełniają wszystkich kryteriów przedstawionych w ramce A, są określane jako „domniemane” neuroprzekaźniki.
Ramka A
Kryteria definiujące neuroprzekaźnik.
Wyróżniające się cechy neuroprzekaźników, w porównaniu z innymi cząsteczkami sygnalizacyjnymi, stają się jaśniejsze przez porównanie z działaniami hormonów wydzielanych przez układ endokrynny. Hormony zazwyczaj wpływają na komórki docelowe znajdujące się daleko od komórki wydzielającej hormon (patrz rozdział 8). To „działanie na odległość” jest osiągane poprzez uwalnianie hormonów do krwiobiegu. W przeciwieństwie do tego, odległość, na jaką działają neuroprzekaźniki, jest niewielka. W wielu synapsach transmitery wiążą się tylko z receptorami na komórce postsynaptycznej, która znajduje się bezpośrednio pod terminalem presynaptycznym (rysunek 6.2A); w takich przypadkach transmiter działa na odległość mniejszą niż mikrometr. Nawet jeśli neuroprzekaźniki rozprzestrzeniają się lokalnie, zmieniając właściwości elektryczne wielu komórek postsynaptycznych (a czasem presynaptycznych) znajdujących się w pobliżu (rysunek 6.2B), działają one jedynie na odległość od kilkudziesięciu do kilkuset mikrometrów. Podczas gdy wydłużone wyrostki aksonalne neuronów pozwalają na uwalnianie neuroprzekaźników nawet w odległości metra od ciała komórki neuronu, przekaźniki te nadal działają tylko w pobliżu presynaptycznego miejsca uwolnienia (rysunek 6.2C).

Rysunek 6.2
Lokalizacja działania neurotransmiterów. Neuroprzekaźniki ogólnie działają albo lokalnie (A), przez zmianę pobudliwości elektrycznej małego obszaru pojedynczej komórki postsynaptycznej, albo bardziej rozproszony (B), przez zmianę pobudliwości elektrycznej (więcej…)
Podczas gdy rozróżnienie między neuroprzekaźnikami i hormonami jest ogólnie jasne, substancja może działać jako neuroprzekaźnik w jednym regionie mózgu, podczas gdy służy jako hormon gdzie indziej. Na przykład, wazopresyna i oksytocyna, dwa hormony peptydowe, które są uwalniane do krążenia z tylnej części przysadki mózgowej, działają również jako neuroprzekaźniki w wielu centralnych synapsach. Szereg innych peptydów również służy zarówno jako hormony, jak i neuroprzekaźniki.