Para melhor compreender este mecanismo, uma simples quebra do caminho do olfato é fornecida abaixo. Quando os humanos mastigam, compostos de sabor volátil são empurrados através do nasofaranx e receptores olfactivos.
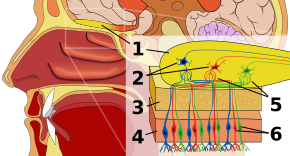
Epitélio olfactivoEdit
A primeira paragem no sistema olfactivo é o epitélio olfactivo, ou tecido que repousa no telhado da cavidade nasal que alberga os receptores olfactivos. Os receptores olfactivos são neurónios bipolares que ligam os odores do ar e se congregam no nervo olfactivo antes de passarem os axónios para os dendritos das células mitrais no bulbo olfactivo. Os receptores sensoriais na boca e nariz são polarizados no estado de repouso, e despolarizam em resposta a alguma mudança no ambiente, como entrar em contato com moléculas odoríferas. As moléculas do odor, que consistem em cadeias de hidrocarbonetos com grupos funcionais, ligam-se aos receptores sensoriais no nariz e na boca. As propriedades dos grupos funcionais incluem: (1) comprimento da cadeia de carbono, (2) grupo terminal, que concordam com diferenças associadas a diferentes odores, (3) grupo lateral, (4) quiralidade, (5) forma, e (6) tamanho. Quando as moléculas odoríferas se ligam aos receptores sensoriais, elas o fazem de acordo com estas propriedades. Cada célula olfativa tem um único tipo de receptor, mas esse receptor pode ser “amplamente sintonizado” e as moléculas odoríferas interagem ainda mais ao nível do receptor, o que significa que, em certos casos, uma molécula odorífera sozinha pode não se ligar a um receptor, mas na presença de outra molécula odorífera, a original se ligaria e assim criaria uma sensação de odor apenas na presença da segunda molécula.
Bolbo olfactivoEditar
No bolbo olfactivo, as moléculas olfactivas são mapeadas espacialmente. Estas representações espaciais são conhecidas como “imagens olfativas”. A representação espacial permite a inibição lateral, ou aumento do contraste e ganho de compressão. O realce do contraste é sensível à mudança e realça os estímulos no cérebro que estão mudando em vez de em repouso. A compressão aumenta a sensibilidade a estímulos de baixa intensidade enquanto diminui a sensibilidade a estímulos de alta intensidade. O bulbo olfativo, embora ainda nos estágios primários de sua compreensão pelos pesquisadores, distingue o olfato dos outros sentidos porque marca um desvio na via sensorial em relação ao que é característico de todos os outros sentidos. Nomeadamente, toda informação sensorial não olfativa passa pelo tálamo após o nível receptor, mas o fato de que a informação do odor, ao invés disso, entra em sua própria área especializada poderia sugerir a história primitiva do olfato e/ou um tipo distinto de processamento da informação do odor no seu caminho para o córtex. O bulbo olfativo abriga glomérulos, ou junções celulares, nos quais convergem milhares de receptores do mesmo tipo, além das células mitrais. Esta organização permite que uma vasta quantidade de informação seja concisamente representada sem a necessidade de um número igualmente grande de tipos de receptores. A combinação resultante da informação do odor é denominada imagem de odor ao nível do bulbo olfativo.
Imaging in the olfactory bulbEdit
2DG methodEdit
Em 1977, o bioquímico Lou Sokoloff, Seymour Kety e Floyd E. Bloom desenvolveram uma forma de mapear a atividade no cérebro através do rastreamento da metabolização do oxigênio pelo cérebro do rato. As células nervosas necessitam de oxigênio e glicose para obter energia. 2-deoxiglicose (2DG) é um isótopo radioativo de glicose que pode ser rastreado no cérebro, pois deixa um traço na célula onde normalmente seria metabolizado por energia se fosse glicose. Após estimulação de uma determinada região de células, fotografias de raios X podem ser cortadas para revelar quais células estavam ativas, particularmente em junções sinápticas.
Ressonância magnética funcional (fMRI) também pode ser usada para medir o metabolismo de um odor. Este método não é terminal como o método da 2-deoxiglicose, assim um animal pode ser medido com muitos odores, e as imagens resultantes podem ser comparadas.
Método da proteína fluorescente verdeEditar
Finalmente, o método da proteína fluorescente verde engendra geneticamente ratos para expressar uma proteína em neurônios ativos, e uma câmera pode então ser colocada dentro do crânio do mouse para medir a atividade.
FindingsEdit
Estes métodos revelam, sobretudo, que a organização da informação olfactiva no bulbo olfactivo é espacial. Padrões moleculares similares resultam em padrões de ativação similares em relação aos glomérulos, e glomérulos que estão mais próximos entre si codificam características similares de informação olfativa.
Córtex olfativoEditar
O córtex olfativo de três camadas, contendo células piramidais, é o próximo ponto de referência no caminho olfativo. Uma célula piramidal recebe informações de uma multiplicidade de células mitrais do bulbo olfativo, fazendo com que o padrão glomerular previamente organizado seja distribuído no córtex olfatório. Esta dispersão de informação das células mitrais permite conexões de feedback auto-excitatório, excitação lateral e auto-inibição e inibição lateral. Estes processos contribuem para o aprendizado Hebbiano, nomeado em homenagem a Donald O. Hebb, e é muitas vezes simplificado pelo ditado “neurônios que disparam juntos fio a fio”. A potencialização a longo prazo, o mecanismo neural para o aprendizado Hebbiano, permite a formação de memória ao nível da célula piramidal. A aprendizagem hebbbica é assim essencialmente o fenômeno pelo qual o córtex olfativo “se lembra” da saída de combinações de moléculas olfativas e permite o reconhecimento de combinações previamente detectadas mais rapidamente do que as novas, combinando-as com a entrada armazenada. Os odores resultantes que anteriormente eram chamados de imagens olfativas são armazenados no córtex olfactivo para reconhecimento são agora referidos como objectos odoríferos. A experiência, portanto, reforça a relação sinal/ruído, na medida em que um objeto de odor previamente detectado pode ser mais facilmente distinguido contra um maior ruído de fundo.
Córtex orbitofrontalEditar
O córtex orbitofrontal (OFC) é o destino final da informação do odor e é onde surge a percepção olfatória consciente. A informação olfactiva entra directamente após a passagem pelo córtex olfactivo, o que marca a distinção de outras informações sensoriais que passam primeiro pelo tálamo. O OFC está localizado dorsal ao córtex pré-frontal, permitindo a entrada direta da informação olfativa no córtex pré-frontal, ou na maior área de decisão do cérebro. Há apenas três tipos de neurônios que cheiram moléculas passam antes de chegar ao OFC: as células receptoras olfatórias no epitélio olfativo, células mitrais e neurônios olfatórios piramidais.
Ao nível do OFC, associações com outras áreas cerebrais são feitas, incluindo entrada da boca (somatossensação), entrada emocional (amígdala), informação visual e informação avaliativa (córtex pré-frontal). O OFC é responsável pelo ajuste seletivo do odor, fusão dos domínios sensoriais e avaliações hedônicas dos odores.