Jotta tätä mekanismia voitaisiin ymmärtää paremmin, seuraavassa esitetään hajupolun yksinkertainen erittely. Kun ihminen pureskelee, haihtuvat makuyhdisteet työntyvät nasofaranxin ja hajureseptorien läpi.
HajuepiteeliMuutos
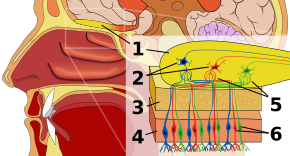
Hajujärjestelmän ensimmäinen pysäkki on hajuepiteeli eli nenäontelon katolla lepäävä kudos, jossa sijaitsevat hajureseptorit. Hajureseptorit ovat bipolaarisia neuroneja, jotka sitovat ilmasta peräisin olevia hajuaineita ja kerääntyvät hajuhermoon ennen kuin aksonit kulkevat hajulohkon mitraalisolujen dendriitteihin. Suun ja nenän aistinvaraiset reseptorit ovat lepotilassa polarisoituneita, ja ne depolarisoituvat vastauksena johonkin ympäristön muutokseen, kuten kosketukseen joutumiseen hajumolekyylien kanssa. Hajumolekyylit, jotka koostuvat hiilivetyketjuista, joissa on funktionaalisia ryhmiä, sitoutuvat nenän ja suun aistireseptoreihin. Funktionaalisten ryhmien ominaisuuksia ovat mm: (1) hiiliketjun pituus, (2) loppuryhmä, joka vastaa eri hajuihin liittyviä eroja, (3) sivuryhmä, (4) kiraalisuus, (5) muoto ja (6) koko. Kun hajumolekyylit sitoutuvat aistireseptoreihin, ne sitoutuvat näiden ominaisuuksien mukaisesti. Jokaisella hajusolulla on yksi reseptorityyppi, mutta reseptori voi olla ”laajasti viritetty”, ja hajumolekyylit ovat edelleen vuorovaikutuksessa reseptoritasolla, mikä tarkoittaa, että tietyissä tapauksissa hajumolekyyli ei välttämättä yksinään sitoudu reseptoriin, mutta toisen hajumolekyylin läsnä ollessa alkuperäinen sitouduisi alkuperäiseen reseptoriin ja näin ollen synnyttäisi hajuaistimuksen vain toisen hajumolekyylin läsnä ollessa.
HajulohkoEdit
Hajulohkossa hajumolekyylit kartoitetaan alueellisesti. Näitä spatiaalisia representaatioita kutsutaan ”hajukuviksi”. Spatiaalinen representaatio mahdollistaa lateraalisen inhibition eli kontrastin vahvistamisen ja vahvistuksen tiivistämisen. Kontrastinvahvistus on herkkä muutoksille ja korostaa aivoissa ärsykkeitä, jotka ovat pikemminkin muuttumassa kuin levossa. Gain compression lisää herkkyyttä matalan intensiteetin ärsykkeille ja vähentää herkkyyttä korkean intensiteetin ärsykkeille. Vaikka tutkijat vielä ymmärtävät hajulohkoa vasta alkuvaiheessa, se erottaa hajun muista aisteista, koska se merkitsee poikkeamaa aistiradassa siitä, mikä on ominaista kaikille muille aisteille. Kaikki muu kuin hajuaistitieto kulkee reseptoritason jälkeen talamuksen kautta, mutta se, että hajutieto sen sijaan kulkee omalle erikoistuneelle alueelleen, voisi viitata hajun primitiiviseen historiaan ja/tai hajutiedon erityyppiseen käsittelyyn matkalla aivokuorelle. Hajulohkossa on glomeruleita eli soluliitoksia, joihin mitraalisolujen lisäksi kokoontuu tuhansia samantyyppisiä reseptoreita. Tämän organisaation ansiosta valtava määrä tietoa voidaan esittää tiiviisti ilman, että tarvitaan yhtä suurta määrää reseptorityyppejä. Tuloksena syntyvää hajuinformaation yhdistelmää kutsutaan hajukuvaksi hajupolvitasolla.
Kuvantaminen hajupolvessaMuokkaa
2DG-menetelmäMuokkaa
Vuonna 1977 biokemistit Lou Sokoloff, Seymour Kety ja Floyd E. Bloom kehittivät tavan, jolla voitiin kartoittaa aivojen toimintaa seuraamalla rotan aivojen hapen metaboliaa. Hermosolut tarvitsevat energiaa varten happea ja glukoosia. 2-deoksiglukoosi (2DG) on radioaktiivinen glukoosi-isotooppi, jota voidaan seurata aivoissa, koska se jättää jäljen soluun, jossa se normaalisti metaboloituisi energiaksi, jos se olisi glukoosia. Kun tiettyä solualuetta on stimuloitu, voidaan röntgenkuvia viipaloimalla paljastaa, mitkä solut olivat aktiivisia, erityisesti synaptisten liitoskohtien kohdalla.
Funktionaalista magneettikuvausta (fMRI) voidaan käyttää myös hajun aineenvaihdunnan mittaamiseen. Tämä menetelmä ei ole terminaalinen kuten 2-deoksiglukoosimenetelmä, joten yhtä eläintä voidaan mitata monilla hajuilla, ja tuloksena saatuja kuvia voidaan verrata.
Vihreän fluoresoivan proteiinin menetelmäMuutos
Viimeiseksi, vihreän fluoresoivan proteiinin menetelmässä hiiret muokataan geneettisesti siten, että ne ilmentävät proteiinia aktiivisissa neuroneissa, minkä jälkeen hiiren kallon sisälle voidaan sijoittaa kamera aktiivisuuden mittaamiseksi.
TuloksetEdit
Nämä menetelmät paljastavat erityisesti sen, että hajutiedon organisoituminen hajupolvessa on avaruudellista. Samankaltaiset molekyylimallit johtavat samankaltaisiin aktivaatiokuvioihin glomerulusten suhteen, ja lähempänä toisiaan olevat glomerulukset koodaavat samankaltaisia hajuinformaation piirteitä.
HajuaivokuoriTiedosto
Kolmekerroksinen, pyramidisoluja sisältävä hajuaivokuori on seuraava vertailukohde hajuradalla. Yksi pyramidisolu vastaanottaa tietoa lukuisista mitraalisoluista hajupolvesta, jolloin aiemmin järjestetty glomerulaarinen kuvio jakautuu hajuaivokuorelle. Tämä mitraalisolujen informaation hajaantuminen mahdollistaa itseä kiihdyttävät takaisinkytkennät, lateraalisen herätteen sekä itse- ja lateraalisen eston. Nämä prosessit edistävät Hebbin oppimista, joka on nimetty Donald O. Hebbin mukaan, ja sitä yksinkertaistetaan usein sanonnalla ”neuronit, jotka palavat yhdessä, johdottuvat yhteen”. Pitkäaikainen potentiaatio, Hebbin oppimisen hermostollinen mekanismi, mahdollistaa muistin muodostumisen pyramidisolujen tasolla. Hebbiläinen oppiminen on siis pohjimmiltaan ilmiö, jonka avulla hajuaivokuori ”muistaa” hajumolekyylien yhdistelmien tuotokset ja mahdollistaa aiemmin aistittujen yhdistelmien tunnistamisen nopeammin kuin uusien yhdistelmien tunnistamisen sovittamalla ne tallennettuihin syötteisiin. Tuloksena syntyviä hajuja, joita aiemmin kutsuttiin hajukuviksi ja jotka tallennetaan hajuaivokuoreen tunnistamista varten, kutsutaan nyt hajuobjekteiksi. Kokemus siis vahvistaa signaali-kohinasuhdetta siten, että aiemmin aistittu hajuobjekti voidaan helpommin erottaa suuremmasta taustakohinasta.
Orbitofrontaalinen aivokuoriMuutos
Orbitofrontaalinen aivokuori (OFC) on hajutiedon lopullinen määränpää, ja se on se paikka, jossa tietoinen hajujen havaitseminen syntyy. Hajutieto tulee suoraan sen jälkeen, kun se on kulkenut hajuaivokuoren läpi, mikä merkitsee eroa muihin aistitietoihin, jotka kulkevat ensin talamuksen läpi. OFC sijaitsee dorsaalisesti prefrontaaliseen aivokuoreen nähden, jolloin hajutieto pääsee suoraan prefrontaaliseen aivokuoreen eli aivojen tärkeimpään päätöksentekoalueeseen. On vain kolme erilaista neuronityyppiä, jotka hajumolekyylit läpäisevät ennen kuin ne saapuvat OFC:hen: hajuepiteelissä olevat hajureseptorisolut, mitraaliset solut ja hajupyramidineuronit.
OfC:n tasolla muodostetaan assosiaatioita muiden aivoalueiden kanssa, mukaan luettuna suusta tuleva tulo (somatosensointi), emotionaalinen tulo (mantelitumake (amygdala), visuaalinen informaatio ja arvioiva informaatio (prefrontaalinen kuori). OFC on vastuussa valikoivasta hajujen virittämisestä, aistialueiden yhdistämisestä ja hajujen hedonisista arvioinneista.