- Introducere
- Rezultate
- Acumularea și atașarea celulelor la minerale necesită gene de chemotaxie și de transport extracelular de electroni în S. oneidensis MR-1
- Caracterizarea congregării în jurul IEA în alte tulpini de Shewanella
- Discuție
- Materiale și metode
- Cultivare și tulpini
- Sinteză minerală
- Experimentele de acumulare a celulelor după fotoblanșire (CAAP) Time-Lapse
- Capturarea prin microscopie a mișcărilor celulelor
- Analiză de înot
- Contribuții ale autorilor
- Declarație privind conflictul de interese
- Recunoștințe
- Supplementary Material
Introducere
La sfârșitul anilor 1980, Shewanella oneidensis MR-1 (Myers și Nealson, 1988a) și mai târziu mai multe specii de Geobacter (Lovley et al., 1993; Champine et al., 2000) s-au dovedit a fi capabile să transfere electroni către acceptori de electroni insolubili (IEA), cum ar fi oxizi metalici insolubili și/sau electrozi încărcați: un proces numit transport extracelular de electroni (EET) (Myers și Nealson, 1988b; Venkateswaran și colab., 1999; Bond și Lovley, 2003). Această capacitate a atras un interes considerabil în ceea ce privește ciclurile biogeochimice, bioremedierea, coroziunea, prelucrarea nanomaterialelor și producția de energie (Bretschger et al., 2007; Kan et al., 2011; Hsu et al., 2012). Deși se știe că mai multe grupuri de microbi sunt capabile de EET, studiile mecaniciste majore au fost realizate doar cu două sisteme model, Shewanella (Fredrickson et al., 2008; Shi et al., 2009) și Geobacter (Lovley et al., 2004).
Până în prezent, mai mult de 100 de alte tulpini de Shewanella au fost izolate dintr-o mare varietate de habitate, inclusiv din coloana de apă deschisă, șisturi de gresie, sedimente marine și de apă dulce, conducte de petrol, saramură de petrol și chiar comunități de alge de pe gheața Antarcticii (Hau și Gralnick, 2007). Genomul a peste 20 dintre aceste specii a fost complet secvențiat (Fredrickson et al., 2008). S-a demonstrat că mai multe dintre aceste specii sunt capabile de EET la IEA, inclusiv S. oneidensis MR-1, Shewanella putrefaciens CN32, S. sp. ANA-3, S. sp. W3-18-1, Shewanella amazonensis SB2B, Shewanella frigidimarina NCIMB 400 și Shewanella loihica PV-4 (Fredrickson et al., 1998; Venkateswaran et al., 1998; Gao et al., 2006; Bretschger, 2008). Alți membri ai genului Shewanella, cum ar fi Shewanella baltica OS217 și Shewanella denitrificans OS155 (tabelul 1), nu sunt capabili de EET (Brettar et al., 2002). Dintre toate aceste tulpini, congregația ca răspuns la IEA a fost studiată doar pentru MR-1.

Tabel 1. Compararea genetică a speciilor de Shewanella și a habitatului lor original.
În cadrul genului Shewanella, mecanismul EET al MR-1 a fost cel mai extensiv caracterizat. MR-1 utilizează mai multe abordări pentru reducerea IEA insolubile: (1) EET directă prin intermediul citocromilor multiheme extracelulare (Beliaev și Saffarini, 1998; Myers și Myers, 2001, 2002; Meyer et al., 2004; Mitchell et al., 2012; Kracke et al., 2015). (Figura 1A); (2) EET mediată utilizând navete electronice solubile legate de citocromi membranari (Lovley et al., 1996; Marsili et al., 2008; Li et al., 2012; Kotloski și Gralnick, 2013; Okamoto et al, 2014); (3) EET mediată care utilizează extensii conductive ale membranei externe care conțin citocromi (Gorby et al., 2006; El-Naggar et al., 2010); și (4) matrici extracelulare conductive care conțin minerale conductoare și semiconductoare (Kato et al., 2010). Mai multe gene au fost identificate în tulpina MR-1 și s-au dovedit a fi esențiale pentru EET (figura 1A și tabelul 2), inclusiv cymA (SO_4591) și combinația mtrBC/omcA (SO_1776, SO_1778 și SO_1779), care codifică componenta decahemă a citocromului c și citocromul c tetrahemă, necesare pentru reducerea mai multor acceptori de electroni anaerobi, inclusiv a oxizilor metalici (Myers și Myers, 2001, 2002; Schwalb et al., 2003). Deoarece toate aceste mecanisme se bazează pe proximitatea celulei față de IEA pentru EET, este important să se înțeleagă detectarea celulelor și migrația netă de înot spre IEA. În ceea ce privește congregația, taxiurile redox sau taxiurile energetice, multe studii au fost efectuate pe MR-1 datorită utilizării versatile a acceptorului de electroni (Bencharit și Ward, 2005; Baraquet et al., 2009; Harris et al., 2010). Taxiul energetic este un termen care cuprinde, în sens larg, aerotaxa, fototaxa, taxiul redox, taxiul către acceptori alternativi de electroni și chimiotaxa către substraturi oxidabile (Alexandre et al., 2004).
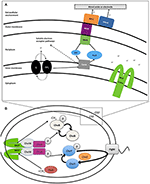
Figura 1. (A) Transportul extracelular de electroni (EET) în Shewanella oneidensis MR-1 construiește forța motrice protonică (pmf). În condiții anaerobe, membrii genului Shewanella pot transporta electroni din membrana internă, periplasma, membrana externă și un electrod prin intermediul unui lanț de citocromi și menaquinone (MQ) cunoscut sub denumirea colectivă de transport extracelular de electroni (EET). Expresia citocromilor EET poate fluctua în funcție de acceptorul de electroni disponibil în mediu. Numărul de grupe heme de tip c care conțin fier este indicat în interiorul (paranteză). Electronii circulă de la donatorii de electroni, cum ar fi lactatul, pentru a reduce chinonele (Q), care transferă simultan H+ în periplasmă, construind pmf, trecând în același timp electroni către CymA. În absența acceptorilor de electroni solubili, electronii sunt transferați către complexul membranei externe MtrCAB. Acest complex MtrCAB poate dona electroni direct către acceptorul terminal de electroni, fie mineral sau electrod anodic, fie prin intermediul moleculelor de flavină. pmf conduce producția de ATP și rotația flagelului polar. Proteinele de chemotaxie care acceptă metil (MCP) „autosensibile” controlează rotația flagelului prin intermediul sistemului de transducție a semnalului de chemotaxie (B) și pot detecta modificări ale concentrației de H+ în timpul reducerii metalelor. (B) Calea probabilă de transducție a semnalului de chemotaxie la Shewanella. Ca răspuns la stimulare, structura MCP se deplasează ca un piston, determinând încetinirea sau oprirea autofosforilării CheA. CheY și CheB nu sunt, prin urmare, fosforilate, iar această lipsă de CheY-P permite o înot lin. Această stimulare are, de asemenea, un alt efect – CheB este inactivă fără fosforilare (nu poate funcționa ca metilesterază) și acest lucru permite proteinei CheR (o metiltransferază activă în permanență) să concureze și să metilizeze în mod liber regiunea de acceptare a metilului din dimerul MCP (domeniul HAMP). Această metilare a MCP acționează pentru a crește rata de autofosforilare a reziduului de histidină CheA. Prin urmare, sistemul de transducție a semnalului are control asupra frecvenței de inversare a flagelului în prezența unor stimuli crescuți sau scăzuți, ceea ce duce la o serie de înot de „alergare și inversare”.

Tabel 2. Genele MR-1 descrise în text.
Este bine documentat faptul că acumularea celulelor MR-1 ca răspuns la acceptorii solubili de electroni este o formă de taxare a energiei, care depinde de fluxul de H+ și de stabilirea unei forțe motrice de protoni (Baraquet et al., 2009) (Figura 1) (Figura 1). O parte a acestui răspuns include o înot mai rapid, așa cum se observă și în cazul unor navete de electroni precum riboflavina sau antrachinona 2,6-disulfonat (AQDS) (Bencharit și Ward, 2005; Harris et al., 2010; Li et al., 2012). În schimb, acumularea de celule în jurul IEA, care a fost numită congregație (Nealson et al., 1995), implică atât creșterea vitezei de înot la contactul cu IEA (numită electrochinezie), cât și creșterea inversării înotului la o scădere a PMF. Inversiunile de înot permit mai multe întâlniri tranzitorii între celule și IEA (cu o durată de 1-100 ms), iar rata la care celulele înotătoare trec la atașarea ireversibilă la IEA în timpul congregației nu a fost încă cuantificată.
Mecanismul (mecanismele) pe care celulele îl (le) utilizează pentru a localiza IEA-urile rămâne neclar (Nealson et al., 1995; Bencharit și Ward, 2005; Harris et al., 2010). Rapoartele timpurii au propus că celălalt organism model studiat, Geobacter, se acumulează în jurul IEA-urilor prin detectarea unui gradient de ioni metalici reduși (Childers et al., 2002), cu toate acestea, ionii metalici reduși nu sunt implicați în mecanismul de detectare utilizat de MR-1 (Bencharit și Ward, 2005). Recent, a fost descrisă congregația bacteriană ca răspuns la electrozi poziționați, ceea ce indică detectarea redox, mai degrabă decât detectarea ionilor metalici, ca declanșator al răspunsului (Harris et al., 2010). Potențialele pozitive aplicate electrodului (200-600 mV față de Ag/AgCl) au determinat congregarea MR-1, similară celei observate în cazul oxizilor metalici (Harris et al., 2012). Potențialele redox ale mineralelor care conțin MnO2 variază între 400 și 600 mV față de Ag/AgCl și acceptă electroni mai ușor decât mineralele Fe(OH)3, care au un potențial echivalent pozat de 100 până la 300 mV față de Ag/AgCl (Burdige, 1993). Într-un studiu anterior, s-a emis ipoteza că înotul caracteristic al MR-1 în jurul IEA este reglementat de doi receptori de chemotaxie cu autosesizare, proteine care acceptă metil (MCP) cu, domeniul receptorului de chemotaxie al canalului Ca+ (Cache) (SO_2240) și domeniul Per/Arnt/Sim (PAS) (SO_1385), și de către proteina kinaza CheA-3 (SO_3207) de transducție a semnalului de chemotaxie, pentru a permite celulei să eșantioneze potențialul redox sau capacitatea de acceptare a electronilor de către o suprafață (tabelul 2 și figura 1B) (Harris et al., 2012). Înainte de acest studiu, nu se cunoșteau genele responsabile de atașarea determinată de motilitate a celulelor pe și în jurul IEA.
În acest raport, studiem mecanismul de congregație a diferitelor tulpini Shewanella prin monitorizarea modelelor de înot și a atașării celulelor la MnO2 și Fe(OH)3. Pe lângă depistarea WT și a mai multor mutanți de deleție ai MR-1, au fost analizate și alte specii de Shewanella (a se vedea tabelul 1). S-a demonstrat că multe tulpini sunt capabile să se congenereze atât în jurul MnO2, cât și în jurul FeOH3, în timp ce altele au răspuns selectiv la MnO2 (SB2B) sau Fe(OH)3 (W3-18-1). Atunci când a fost monitorizată atașarea celulelor la suprafețele minerale, am observat o corelație puternică între capacitatea celulei de a se congrega și atașarea celulelor la suprafața IEA. Am comparat apoi genomurile acestor specii pentru a găsi genele candidate implicate în înotul de congregație, acumularea și fenotipurile de atașare a celulelor ca răspuns la suprafețe IEA specifice.
Rezultate
Acumularea și atașarea celulelor la minerale necesită gene de chemotaxie și de transport extracelular de electroni în S. oneidensis MR-1
Majoritatea studiilor timpurii privind taxiul energetic în MR-1 au utilizat o metodă de testare a plăcilor de roi (Nealson et al., 1995; Baraquet et al., 2009; Li et al., 2012). Aici, oferim o caracterizare mai aprofundată a acestor mecanisme de taxare a energiei încă necunoscute prin utilizarea unei metode cantitative de urmărire a înotului celulelor individuale prin medii lichide ca răspuns la IEA și a acumulării de celule după fotoblanșire (CAAP) pentru a măsura rata de atașare a celulelor (Li et al., 2010). CAAP utilizează iradierea cu laser UV de la un microscop confocal pentru a întuneca ireversibil (stinge fluorescența) celulele marcate cu GFP pe și în jurul unei anumite IEA. Prin cuantificarea celulelor fluorescente pe măsură ce se deplasează în zona întunecată, pot fi cuantificate ratele de acumulare și de atașare și pot fi comparate diferite tulpini. Pentru a fi considerată atașată, poziția celulei mobile este verificată prin analizarea unei imagini 3D a câmpului vizual. În cazul în care celula ajunge să se odihnească pe suprafața minerală, atunci celula este considerată „atașată”. Rezistența legăturilor dintre celulele MR-1 staționare și suprafețe a fost măsurată cu ajutorul unor pensete optice într-un studiu separat (Gross și El-Naggar, 2015). Această metodă permite distincția între atașarea dirijată a celulelor și atașarea electrostatică aleatorie.
După cum se arată în figura 2, celulele WT MR-1 s-au atașat la suprafața minerală după 30 de minute, cu o oarecare abatere mare în ceea ce privește numărul, 2.655 ± 1.352 celule/mm2. În acest timp, celulele prezintă un înot de tip „touch and go”, realizând un contact tranzitoriu cu suprafața minerală MnO2. După 2 h, numărul de celule noi atașate la suprafața minerală a crescut la 4 300 ± 584 de celule/mm2 de suprafață minerală, în timp ce celulele motile suplimentare au continuat să se adune (Videoclipurile S1, S2 și S17). Mutanții cu deleții triple în genele cheie EET mtrBC/omcA (SO_1776, SO_1778, SO_1779), deleția unică a cymA (SO_4591) și chemoreceptorul principal de taxare a energiei MCP Cache (SO_2240) au fost legați anterior de fenotipul de congregație a înotului (Harris et al., 2012). Mutanții de deleție (ΔcymA, Δmcp_cache și ΔmtrBC/omcA) au fost cu toții mobili și capabili să inverseze direcția de înot, dar au fost incapabili de congregație și au arătat puțin sau deloc atașament la MnO2 în timpul experimentului (Figura 2C și Figurile S1A,B). Acumularea WT în zona decolorată la t = 2 h este în medie de 4.300 ± 584 celule/mm2 , în timp ce mutanții Δmcp_cache, și ΔmtrBC/omcA se acumulează în număr neglijabil (Figura 2C și S1CD).
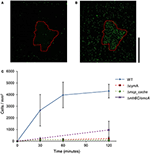
Figura 2. MR-1 înotător poate migra spre minerale insolubile acceptoare de electroni și se poate atașa. Imagine reprezentativă de microscopie confocală cu fluorescență a celulelor WT MR-1 la t = 0 și t = 120 min (A,B). Celulele WT MR-1 GFP care înoată WT MR-1 au fost introduse în particule de MnO2 (contur punctat roșu) în capilarul sigilat anaerob. La t = 0, toate celulele au fost fotodecolorate ireversibil într-o zonă de 250 pe 250 μm în jurul particulei (A). Apoi au fost numărate celulele fluorescente din afara zonei decolorate care înoată în cadru și se atașează la suprafața minerală. Bara de scară verticală neagră din dreapta reprezintă 100 μm. Graficul (C) compară atașamentul WT MR-1 pe particula de MnO2 pe parcursul a 120 de minute cu chemotaxia și mutanții de deleție a transferului extracelular de electroni (MR-1 Δmcp_cache, ΔcymA și ΔmtrBC/omcA). Barele de eroare includ 2 deviații standard.
Caracterizarea congregării în jurul IEA în alte tulpini de Shewanella
Nouă tulpini de Shewanella au fost testate pentru capacitatea lor de a se congrega în jurul suprafețelor minerale de MnO2 și Fe(OH)3 cu ajutorul unor serii de timp de testare a atașării celulelor la mineral (tabelul 3 și videoclipurile S3-S14). Ca și în cazul MR-1, toate cele nouă tulpini examinate aveau un singur flagel polar, iar inversarea direcției de înot a fost realizată prin inversarea rotației flagelare (datele nu sunt prezentate). Urmele de înot din cadrul aceluiași experiment au fost sortate în două grupuri separate pe baza traiectoriei de înot (Tabelul 3): cele care au intrat în contact cu suprafața insolubilă de oxid metalic (au înotat în limita a 2 μm), comparativ cu cele care nu au intrat în contact (au înotat >2 μm). Înotătorii care au intrat în contact și care au demonstrat o creștere semnificativă a frecvenței de inversare și a vitezei de înot față de grupul care nu a intrat în contact (P < 0,05) sunt clasificați ca fiind pozitivi pentru comportamentul de congregație, așa cum sunt desemnați cu litere superscript (Tabelul 3).

Tabel 3. Viteza de înot a bacteriilor și frecvența de inversare în jurul mineralelor de oxizi metalici.
Sursele ANA3, CN32 și PV-4, în plus față de MR-1, au fost pozitive pentru congregație atât în jurul MnO2, cât și în jurul Fe(OH)3 (Tabelul 3, Figura 3; Videoclipuri S1-S8). Acestea au prezentat rate maxime de inversare atunci când au fost situate între 5 și 40 μm de la suprafața particulelor de MnO2 sau FeOH3 (Figura 3). Trei tulpini: OS217, NCIMB400 și OS155 nu au fost motile la timpul t = 30 min – ca răspuns la mineralele Fe(OH)3 sau MnO2 (tabelul 3). De exemplu, frecvența de inversare a celulelor MR-1 înotătoare care au intrat în contact cu MnO2 a fost de 0,94 ± 0,53 inversări/s, în timp ce frecvența de inversare a celulelor care nu au intrat în contact a fost de 0,62 ± 0,73 inversări/s. Viteza a crescut, de asemenea, în grupul de MR-1 în contact, de la 19,26 ± 11,2 μm/s în grupul fără contact la 24,37 ± 6 μm/s în grupul în contact. Deoarece a existat atât o creștere semnificativă din punct de vedere statistic a vitezei de înot, cât și inversări pentru a permite celulelor să rămână în apropierea particulei de oxid metalic, s-a spus că această tulpină a fost congregație pozitivă.
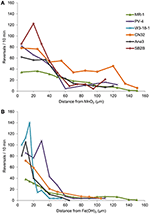
Figura 3. (A,B) Frecvența de inversare (axa y) a celulelor Shewanella care înoată în funcție de distanța (axa x) față de particula IEA. Frecvențele medii de inversare ale celulelor înotătoare urmărite individual sunt împărțite în intervale de 5 μm de-a lungul axei x pentru experimentele cu particule de MnO2 (A) sau Fe(OH)3 (B).
Pistele de înot ale Shewanella spp. (30 s) care au demonstrat o preferință pentru mineralele de oxizi metalici sunt evidențiate în figurile 4A-F. Figura SB2B celulele nu au prezentat niciun răspuns de înot la Fe(OH)3 (figura 4D), în timp ce au prezentat o congregație activă în jurul particulelor de MnO2 (figura 4A). În schimb, celulele W3-18-1 s-au adunat în jurul particulelor de Fe(OH)3 (Figura 4C), dar au prezentat o activitate de înot și de inversare semnificativ diminuată în jurul MnO2 (Figura 4F; Videoclipurile S9, S14).
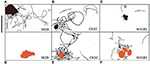
Figura 4. (A-F) Panou de trei trasee de înot ale Shewanella spp. (30 s) a demonstrat o preferință pentru mineralele de oxizi metalici. A fost studiat comportamentul de înot a trei Shewanella spp. motile ca răspuns la MnO2 (rândul de sus – particulă colorată în negru) și Fe(OH)3 (rândul de jos – particulă colorată în portocaliu). Congregația are loc în condiții anaerobe, iar urmele de înot sunt reprezentate cu negru. SB2B (stânga), CN32 (centru) și W3181 (dreapta). Înotul celulelor a fost urmărit manual. Bara de scară din stânga sus = 50 μm. Forma de culoare maro reprezintă MnO2, iar cea portocalie reprezintă Fe(OH)3. Atunci când nu sunt detectate celule mobile, acest lucru este indicat prin absența liniilor negre.
Discuție
Membrii genului Shewanella sunt cuprinși de aerobi heterotrofi, facultativi, capabili să utilizeze o gamă largă de substanțe organice și anorganice ca surse de energie. Numeroase tulpini de Shewanella au fost izolate din coloana de apă și din habitatele sedimentare în locații din întreaga lume. În plus față de acceptorii de electroni solubili, multe dintre aceste organisme pot respira o gamă largă de oxizi metalici insolubili, care apar în mod natural, în condiții anoxice.
În studiul nostru privind comportamentul de congregație la opt specii de Shewanella, am folosit urmărirea celulelor cu ajutorul analizei computerizate și testarea seriilor temporale de atașare a celulelor la MnO2 sau Fe(OH)3 pentru a demonstra că cinci din cele opt shewanellae sunt capabile de acest comportament CN32, ANA-3, W3-18-1, SB2B și PV-4) și că unele tulpini (W3-18-1, SB2B) prezintă o preferință pentru un oxid de metal în detrimentul celuilalt în aceste condiții (figura 4). Astfel de constatări sunt în concordanță cu ideea că aceste bacterii s-au adaptat la acceptorul de electroni insolubil predominant găsit în habitatul din care au fost izolate. În plus, trei tulpini Shewanella care au fost izolate din coloana de apă nu s-au adunat ca răspuns la IEA. Dacă aceste diferențe derivă din absența genelor implicate în EET (ca în cazul OS217) sau din alte motive va fi un punct pentru studii viitoare.
În alte studii, creșterea peste noapte a tulpinilor de Shewanella care compară ratele de reducere a diferiților oxizi metalici în comparație cap la cap, s-a demonstrat că reduc în mod preferențial oxizii metalici din nișa lor de mediu (Bretschger et al., 2007). Rezultatele prezentate aici sunt în concordanță cu ipoteza conform căreia speciile Shewanella au evoluat un comportament de congregație și de atașare în concordanță cu mediile din care au fost izolate (tabelul 3). De exemplu, W3-18-1 pare să dezvăluie o înclinație semnificativă pentru congregație în prezența și atașarea la minerale Fe(OH)3 native din platoul continental al Pacificului (Harris et al., 2012), în timp ce SB2B preferă MnO2 (Venkateswaran et al., 1998), similar cu cel din sedimentele fluviului Amazon.
Compararea genomică din tabelul 1 sugerează că prezența citocromului octaheme (SO_4142) poate fi importantă pentru înotul ca răspuns la potențialele redox relativ „scăzute” (100-300 mV față de Ag/AgCl) ale Fe(OH)3. În timp ce mtrF (SO_1780) poate fi necesar pentru răspunsul la potențialele redox relativ „mai mari” ale MnO2 (între 400 și 600 mV față de Ag/AgCl). Această ipoteză ar putea fi apoi testată cu mutanți de deleție în MR-1. Această lucrare completează lucrările anterioare ale lui Harris et al. (2012), dezvăluind că genele implicate în efectul net al comportamentului de motilitate față de minerale IEA sau electrozi este cumulativ prin scări temporale relevante de 1-30 min (Figura 2) (Harris et al., 2012). Constatările noastre cu ajutorul tehnicii de microscopie confocală CAAP, au demonstrat că congregația duce la o migrare lentă a celulelor spre mineralele IEA cu atașare eventuală. Astfel, acest comportament este denumit în mod corespunzător congregație, deoarece descrie adunarea sau acumularea de bacterii mobile în jurul IEA. Am arătat aici că genele mcp cache și mtrBC/omcA sunt esențiale pentru fenotipul de acumulare și atașare la MR-1, pe lângă faptul că sunt responsabile pentru modelele de înot caracteristice de viteză crescută și comportament de tip „run-and-reversal” care a fost identificat anterior (Harris et al., 2012).
Rezultatele experimentelor cu mutanții ΔcymA, ΔmtrBC/omcA și Δmcp_cache în MR-1 arată o inhibiție a comportamentului de acumulare, atașare și congregație ca răspuns la toate IEA. Prin urmare, se poate emite ipoteza că prezența genelor EET omoloage (cymA, mtrB, mtrC, omcA) și a genei de chemotaxie acceptoare de metil (mcp_cache) determină răspunsurile fenotipice pe care le observăm la alte Shewanella spp. (tabelul 3). Comparația genetică a tulpinilor, care ar putea răspunde la IEA cu potențial redox „inferior”, indică faptul că citocromii octahemici periferici din membrana externă (cum ar fi SO_4142) pot juca un anumit rol în răspunsul la IEA cu potențial redox „inferior”. Prin urmare, aceste observații privind înotul susținut în jurul unor minerale specifice corespund unor genotipuri specifice ale diferitelor specii.
În timpul multor experimente de MFC (Kotloski și Gralnick, 2013) și a experimentelor de cultură pe loturi cu reducere a metalelor, nu a fost măsurat taxiul de energie mediat de transferul de electroni extracelular, sau capacitatea de congregație (Kotloski și Gralnick, 2013). În cadrul testelor de reducere a oxizilor metalici, distribuția 3D a celulelor și motilitatea celulară ar putea influența în mare măsură reducerea metalelor din cauza vitezei de agitare a incubatorului, a dimensiunilor flacoanelor de cultură sau a amestecului. Comportamentul de congregație influențează în mod direct atașarea celulelor la IEA. Relația dintre congregație și formarea apendicelui de tip nanofire este încă necunoscută (Pirbadian et al., 2015), deși parametrii de motilitate și congregație sunt rareori monitorizați în aceste studii (Gorby et al., 2005; Reguera et al., 2005). Transformarea acestor gene de congregație de bază (cymA, mtrB, mtrA, omcA, mcp_pas și mcp_cache) în alte specii de bacterii, cu un singur flagel polar, ar putea, într-o bună zi, să îmbunătățească capacitățile de bioremediere, putând induce atașarea și colonizarea bacteriană a unor suprafețe care, altfel, ar fi dificile sau imposibile.
Materiale și metode
Cultivare și tulpini
MR-1 și mai mulți mutanți de deleție proveniți din MR-1 au fost examinați în acest studiu (tabelul 2) (Beliaev și Saffarini, 1998; Myers și Myers, 2002). Tulpinile au fost inoculate din stocurile de glicerină păstrate la -80°C pe plăci Luria-Bertani (LB) și cultivate peste noapte la 30°C. Coloniile individuale au fost apoi selectate și inoculate în 5 ml de mediu minimal definit (M1) (Bretschger et al., 2007) suplimentat cu 18 mM de lactat ca sursă de energie (Bretschger et al., 2007) în tuburi de 15 ml (VWR International LLC, Randor, Pennsylvania, SUA) și incubate orizontal într-un agitator (180 rpm) timp de 48 de ore la 30 °C. Densitatea optică a fost măsurată cu ajutorul unui spectrofotometru (Unico 1100RS spectrofotometru, Dayton, New Jersey, SUA). Celulele au fost prelevate la o OD600 de 0,5 ± 0,2 (după ~48 h). În experimentele de înot, culturile de cinci mililitri au fost prelevate atunci când celulele au atins o DO de 0,4, au fost amestecate cu oxizi de mangan sau de fier și au fost introduse într-un capilar de sticlă (0,02 × 0,20 mm) (Vitrocom, Mountain Lakes, New Jersey, SUA), care a fost apoi sigilat cu ajutorul unei grăsimi de vid, așa cum a fost descris anterior (Harris et al, 2010).
Sinteză minerală
Soluția stoc de Fe(OH)3 a fost preparată în conformitate cu protocolul lui Cornell și Schwertmann și apoi verificată prin defracție cu raze X (Schwertmann și Cornell, 2008). Acest preparat de MnO2 coloidal a început cu 8 g de KMnO4 dizolvat în 200 ml, utilizând în același timp toate măsurile de siguranță posibile. Soluția a fost amestecată continuu folosind o bară de agitare magnetică la nivel înalt și încălzită sub temperatura de fierbere. Apoi, s-au adăugat 5 mL de hidroxid de sodiu 10 M pentru a neutraliza acidul produs de reacție. Într-un balon separat, 15 g de clorură de mangan au fost dizolvate în 75 ml de apă distilată. În cele din urmă, soluția a fost apoi amestecată lent cu soluția de permanganat (într-o hotă chimică) timp de 75 min. După ce s-a răcit soluția, aceasta a fost apoi spălată prin centrifugare și clătită cu apă deionizată (DI) (18 Meg-Ohm cm) de cinci ori. Precipitatul final a fost lăsat să se usuce prin filtrare în vid într-o bancă curată și a fost desecat timp de 36 h. Mineralele rezultate au fost analizate prin difracție de raze X pentru a confirma producerea de Fe(OH)3 și MnO2 (Bretschger et al., 2007; Salas et al., 2010).
Particulele minerale suspendate au fost amestecate cu cultura la o concentrație finală de 300 mg/mL de MnO2 sau Fe(OH)3. Celulele au fost apoi scufundate prin capilaritate în tuburi capilare dreptunghiulare (0,02 × 0,20 mm) (Vitrocom, Mountain Lakes, New Jersey, SUA). Tuburile au fost sigilate cu unsoare de silicon în vid (Dow Corning, Midland, Milwaukie, SUA) și au fost observate cu ajutorul microscopiei luminoase, microscopiei cu fluorescență și microscopiei confocale.
Experimentele de acumulare a celulelor după fotoblanșire (CAAP) Time-Lapse
Toate tulpinile marcate fluorescent (GFP) au fost transformate așa cum a fost descris anterior (McLean et al, 2008) și apoi cultivate aerobic pe un mediu minimal definit cu 25 μg/ml de kanamicină și 18 mM de lactat timp de 48 h la 30°C. Culturile de cinci mililitri au fost prelevate atunci când celulele au atins o DO de 0,4, au fost amestecate cu oxizi de mangan sau de fier și introduse într-un capilar de sticlă (0,02 × 0,20 mm) (Vitrocom, Mountain Lakes, New Jersey, SUA), care a fost apoi sigilat cu ajutorul unei grăsimi de vid, așa cum a fost descris anterior (Harris et al., 2010). Celulele WT MR-1, ΔcymA, Δmcp_cache, Δmcp_cache și ΔmtrBC/omcA marcate cu GFP au fost albite folosind setările de intensitate maximă a luminii cu lentile 60X, ale unui microscop confocal inversat Leica TCS SPE (Wetzlar, Germania) timp de 15 min. Pentru a se asigura că a avut loc decolorarea, time-lapse a capturat o zonă de ecran (zonă selectată de 250 × 250 μm) la fiecare minut până când celulele originale au apărut întunecate, iar celulele din jur au rămas puternic fluorescente. Imaginile au fost apoi capturate folosind o excitație de 588 nm și o emisie de 530 nm. O înregistrare video time-lapse a întregii secțiuni a tubului a fost capturată cu ajutorul software-ului Leica Imaging și a funcției „autofocus” pentru următoarele 3 h. Celulele au fost, de asemenea, observate în modul de lumină transmisă pentru a verifica dacă celulele albite erau mobile și intacte. Un control negativ separat, cu Δmcp_cache marcat cu GFP, a fost capturat timp de 3 h. În acest control negativ nu au fost observate celule care să se acumuleze în zona întunecată și nici celulele decolorate nu au recuperat fluorescența GFP. Răspunsul întregului capilar (înălțime de 30 μm) a fost capturat cu ajutorul fotografiei time-lapse, iar suma celulelor din toate cele 20 de stive pe axa z a fost determinată (folosind metoda de analiză computerizată de mai jos) pentru fiecare punct de timp.
Capturarea prin microscopie a mișcărilor celulelor
Metodele de urmărire și analiză a bacteriilor au fost identice cu studiile anterioare utilizate pentru Figura 3 și înregistrate în Tabelul 3 (Harris et al., 2012). Atât urmele de înot bacteriene computerizate, cât și cele manuale au fost standardizate folosind o riglă de scară (100 μm). Din fiecare experiment, activitatea globală de înot în cadrul cadrului video – echivalent cu un câmp vizual de 107 × 193 μm – a fost înregistrată și videoclipul a fost normalizat în timp pentru a oferi viteze de înot în μm/s. Mai mulți parametri au fost măsurați pentru fiecare urmă de înot bacteriană, cum ar fi distanța totală parcursă, timpul de urmărire de la prima apariție și dispariție a bacteriilor, numărul de inversări, distanța dintre fiecare inversare și oxidul metalic și distanța dintre oxidul metalic și începutul urmei bacteriene.
Analiză de înot
Celulele au fost urmărite manual din datele video (30 de cadre/s), cadru cu cadru, și măsurate cu ajutorul software-ului de analiză a imaginilor ImageJ. Datele introduse includ viteza de înot, poziția de pornire a bacteriilor în raport cu cea mai apropiată suprafață IEA și poziția fiecărui eveniment de inversare bacteriană a fost identificată și înregistrată în raport cu distanța față de cea mai apropiată suprafață IEA (figura 3 și tabelul 3). Pentru determinarea activității de înot după contactul cu IEA, celulele înotătoare au fost împărțite în două grupuri pentru analiză: celulele care au înotat la mai puțin de 2 μm de o particulă au fost considerate „în contact”, iar cele care nu au înotat la mai puțin de 2 μm de suprafața particulei au fost considerate „fără contact”. În plus față de metodele de urmărire manuală descrise mai sus, datele experimentale au fost apoi sortate pe calculator și analizate cu ajutorul unui algoritm descris anterior pentru a obține datele de înot calculate (Crocker și Grier, 1996; Harris et al., 2012; Harris, 2013). Pentru a produce figurile 3, 4, toate datele experimentale din patru replici biologice separate au fost combinate, puse în comun și mediate în intervale în funcție de distanța față de particula IEA. Deoarece aceste date au fost grupate în distanțe, barele de eroare se suprapun; nu se poate face nicio comparație a frecvenței de inversare, la o anumită distanță, între tulpini, deoarece această metodă de vizualizare exclude identificatorii de experimente.
Contribuții ale autorilor
HH: a proiectat și a efectuat experimentele; HH și IS-A: au scris și revizuit lucrarea; ES, ME-N și JM: au ajutat la proiectarea experimentelor; WT: a ajutat la efectuarea analizei datelor; JM și KN: au revizuit manuscrisul.
Declarație privind conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Recunoștințe
Mulțumiri speciale lui Mandy J. Ward și Gijs Kuenen pentru sfaturi privind cercetarea și proiectarea experimentelor. Îi mulțumim lui Meaghan Sullivan pentru analizele manuale de urmărire. Le mulțumim lui Cecile Jourlin-Castelli, Samantha Reed, Jun Li și David Culley pentru furnizarea mutanților Δmcp_cache, ΔmtrBC/omcA și ΔcymA. Finanțarea pentru această lucrare a fost asigurată de un premiu al Air Force Office of Scientific Research Award . Cercetarea IS-A a fost susținută de bursa NASA Planetary Biology Internship.
Supplementary Material
Materialul suplimentar pentru acest articol poate fi găsit online la adresa: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02568/full#supplementary-material
Alexandre, G., Greer-Phillips, S., și Zhulin, I. B. (2004). Rolul ecologic al taxiurilor energetice în microorganisme. FEMS Microbiol. Rev. 28, 113-126. doi: 10.1016/j.femsre.2003.10.003
PubMed Abstract | Textul integral | Google Scholar
Baraquet, C., Théraulaz, L., Iobbi-Nivol, C., Méjean, V., și Jourlin-Castelli, C. (2009). Chemoreceptorii neașteptați mediază taxarea energiei către acceptorii de electroni în Shewanella oneidensis. Mol. Microbiol. 73, 278-290. doi: 10.1111/j.1365-2958.2009.06770.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Beliaev, A. S., and Saffarini, D. A. (1998). Shewanella putrefaciens mtrB codifică o proteină a membranei externe necesară pentru reducerea Fe(III) și Mn(IV). J. Bacteriol. 180, 6292-6297.
PubMed Abstract | Google Scholar
Bencharit, S., și Ward, M. J. (2005). Răspunsuri chemotactice la metale și acceptori de electroni anaerobi în Shewanella oneidensis MR-1. J. Bacteriol. 187, 5049-5053. doi: 10.1128/JB.187.14.5049-5053.2005
PubMed Abstract | CrossRef Full Text | Google Scholar
Bond, D. R., și Lovley, D. R. (2003). Producerea de energie electrică de către Geobacter sulfurreducens atașat la electrozi. Appl. Environ. Microbiol. 69, 1548-1555. doi: 10.1128/AEM.69.3.1548-1555.2003
PubMed Abstract | CrossRef Full Text | Google Scholar
Bowman, J. P., McCammon, S. A., Nichols, D. S. S., Skerratt, J. H., Rea, S. M., Nichols, P. D., et al. (1997). Shewanella gelidimarina sp. nov. și Shewanella frigidimarina sp. nov., noi specii antarctice cu capacitatea de a produce acid eicosapentaenoic (20:5ω3) și de a se dezvolta în condiții anaerobe prin reducerea disimilatorie a Fe(III). Int. J. Syst. Bacteriol. 47, 1040-1047. doi: 10.1099/00207713-47-4-1040
PubMed Abstract | Text integral | Google Scholar
Bretschger, O. (2008). Electron Transfer Capability and Metabolic Processes of the Genus Shewanella with Applications to the Optimization of Microbial Fuel Cells. University of Southern California, ProQuest Dissertations Publishing, 3325185.
Google Scholar
Bretschger, O., Obraztsova, A., Sturm, C. A., Chang, I. S., Gorby, Y. A., Reed, S. B., et al. (2007). Current production and metal oxide reduction by Shewanella oneidensis MR-1 wild type and mutants. Appl. Environ. Microbiol. 73:7003. doi: 10.1128/AEM.01087-07
PubMed Abstract | Refef Full Text | Google Scholar
Brettar, I., Christen, R., and Höfle, M. G. (2002). Shewanella denitrificans sp nov., o bacterie denitrificatoare viguroasă izolată din interfața oxic-anoxică a adâncului Gotland din Marea Baltică centrală. Int. J. Syst. Evol. microbiol. 52, 2211-2217. doi: 10.1099/00207713-52-6-2211
PubMed Abstract | Full CrossRef Text | Google Scholar
Burdige, D. J. (1993). Biogeochimia reducerii manganului și a fierului în sedimentele marine. Earth Sci. Rev. 35, 249-284. doi: 10.1016/0012-8252(93)90040-E
CrossRef Full Text | Google Scholar
Champine, J. E., Underhill, B., Johnston, J. M., Lilly, W. W., și Goodwin, S. (2000). Electron transfer in the dissimilatory iron-reducing bacterium Geobacter metallireducens. Anaerobe 6, 187-196. doi: 10.1006/anae.2000.0333
CrossRef Full Text | Google Scholar
Childers, S. E., Ciufo, S., and Lovley, D. R. (2002). Geobacter metallireducens accesează oxidul insolubil de Fe(III) prin chemotaxie. Nature 416, 767-769. doi: 10.1038/416767a
PubMed Abstract | Text integral | Google Scholar
Coursolle, D., și Gralnick, J. A. (2010). Modularitatea căii respiratorii Mtr a Shewanella oneidensis tulpina MR-1. Mol. Microbiol. 77, 995-1008. doi: 10.1111/j.1365-2958.2010.07266.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Crocker, J. C., și Grier, D. G. (1996). Metode de microscopie video digitală pentru studii coloidale. J. Colloid Interface Sci. 179, 298-310. doi: 10.1006/jcis.1996.0217
CrossRef Full Text | Google Scholar
El-Naggar, M. Y., Wanger, G., Leung, K. M., Yuzvinsky, T. D., Southam, G., Yang, J., et al. (2010). Transportul electric de-a lungul nanofirelor bacteriene din Shewanella oneidensis MR-1. Proc. Natl Acad. Sci. U.S.A. 107, 18127-18131. doi: 10.1073/pnas.1004880107
PubMed Abstract | CrossRef Full Text | Google Scholar
Fredrickson, J. K., Romine, M. F., Beliaev, A. S., Auchtung, J. M., Driscoll, M. E., Gardner, T. S., et al. (2008). Towards environmental systems biology of Shewanella. Nat. Rev. Microbiol. 6, 592-603. doi: 10.1038/nrmicro1947
PubMed Abstract | Text integral | Google Scholar
Fredrickson, J. K., Zachara, J. M., Kennedy, D. W., Dong, H., Onstott, T. C., Hinman, N. W., et al. (1998). Biogenic iron mineralization accompanying the dissimilatory reduction of hydrous ferric oxide by a groundwater bacterium. Geochim. Cosmochim. Acta 62, 3239-3257. doi: 10.1016/S0016-7037(98)00243-9
CrossRef Full Text | Google Scholar
Gao, H., Obraztova, A., Stewart, N., Popa, R., Fredrickson, J. K., Tiedje, J. M., et al. (2006). Shewanella loihica sp. nov., izolată din covoarele microbiene bogate în fier din Oceanul Pacific. Int. J. Syst. Evol. microbiol. 56, 1911-1916. doi: 10.1099/ijs.0.64354-0
PubMed Abstract | Text integral | Google Scholar
Gorby, Y. A., Beveridge, T. J., și Wiley, W. R. (2005). „Composition, reactivity, and regulation of extracellular metal-reducing structures (nanowires) produced by dissimilatory metal reducing bacteria,” în Annual NABIR PI Meeting (Warrenton, VA).
Google Scholar
Gorby, Y. A., Yanina, S., McLean, J. S., Rosso, K. M., Moyles, D., Dohnalkova, A., et al. (2006). Nanofire bacteriene conductoare de electricitate produse de Shewanella oneidensis tulpina MR-1 și de alte microorganisme. Proc. Natl. acad. Sci. U.S.A. 103, 11358-11363. doi: 10.1073/pnas.0604517103
PubMed Abstract | CrossRef Full Text | Google Scholar
Gross, B. J., și El-Naggar, M. Y. (2015). O platformă combinată de captare electrochimică și optică pentru măsurarea ratelor de respirație a celulelor unice la interfețele electrozilor. Rev. Sci. Instrum. Instrum. 86:064301. doi: 10.1063/1.4922853
PubMed Abstract | Textul integral | Google Scholar
Harris, H. W. (2013). Identificarea unui nou mecanism de detecție bacteriană: Characterization of Bacterial Insoluble Electron Acceptor Sensing. University of Southern California.
Google Scholar
Harris, H. W., El-Naggar, M. Y., Bretschger, O., Ward, M. J., Romine, M. F., Obraztsova, A. Y., et al. (2010). Electrokinezia este un comportament microbian care necesită transportul extracelular de electroni. Proc. Natl. acad. Sci. U.S.A. 107, 326-331. doi: 10.1073/pnas.0907468107
PubMed Abstract | CrossRef Full Text | Google Scholar
Harris, H. W., Naggar, M. Y., și Nealson, K. H. (2012). Shewanella oneidensis MR-1 chemotaxis proteins and electron-transport chain components essential for congregation near insoluble electron acceptors. Biochem. Soc. Trans. 40, 1167-1177. doi: 10.1042/BST20120232
PubMed Abstract | Textul integral | Google Scholar
Hau, H. H. H., și Gralnick, J. A. (2007). Ecologia și biotehnologia genului Shewanella. Annu. Rev. Microbiol. 61, 237-258. doi: 10.1146/annurev.micro.61.080706.093257
PubMed Abstract | CrossRef Full Text | Google Scholar
Hsu, L., Masuda, S. A., Nealson, K. H., și Pirbazari, M. (2012). Evaluarea biocatozilor Shewanella cu celule de combustibil microbian pentru tratarea contaminării cu cromat. R. Soc. Chem. Adv. 2, 5844-5855. doi: 10.1039/c2ra20478a
CrossRef Full Text | Google Scholar
Kan, J., Hsu, L., Cheung, A. C. M., Pirbazari, M., și Nealson, K. H. (2011). Producția de curent de către comunitățile bacteriene în celule de combustibil microbiene îmbogățite din nămol de apă uzată cu diferiți donatori de electroni. Environ. Sci. Technol. 45, 1139-1146. doi: 10.1021/es102645v
PubMed Abstract | Text integral | Google Scholar
Kato, S., Nakamura, R., Kai, F., Watanabe, K., și Hashimoto, K. (2010). Interacțiuni respiratorii ale bacteriilor din sol cu minerale (semi)conductoare de oxid de fier. Environ. Microbiol. 12, 3114-3123. doi: 10.1111/j.1462-2920.2010.02284.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Kotloski, N. J., și Gralnick, J. A. (2013). Navetele electronice de flavin domină transferul extracelular de electroni de către Shewanella oneidensis. MBio 4:e00553. doi: 10.1128/mBio.00553-12
PubMed Abstract | Full CrossRef Text | Google Scholar
Kracke, F., Vassilev, I. și Krömer, J. O. (2015). Transportul microbian de electroni și conservarea energiei – fundamentul pentru optimizarea sistemelor bioelectrochimice. Front. Microbiol. 6:575. doi: 10.3389/fmicb.2015.00575
PubMed Abstract | Textul integral | Google Scholar
Li, J., Go, A. C., Ward, M. J., și Ottemann, K. M. (2010). The chemical-in-plug bacterial chemotaxis assay is prone to false positive responses. BMC Res. Notes 3:77. doi: 10.1186/1756-0500-3-77
PubMed Abstract | Text integral | Google Scholar
Li, J., Romine, M. F., și Ward, M. J. (2007). Identificarea și analiza unui grup de gene de chemotaxis foarte conservate în speciile Shewanella. FEMS Microbiol. Lett. 273, 180-186. doi: 10.1111/j.1574-6968.2007.00810.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Li, R., Tiedje, J. M., Chiu, C., și Worden, R. M. (2012). Navetele electronice solubile pot media taxiul de energie către acceptorii de electroni insolubili. Environ. Sci. Technol. 46, 2813-2820. doi: 10.1021/es204302w
PubMed Abstract | Text integral | Google Scholar
Lovley, D. R., Coates, J. D., Blunt-Harris, E. L., Phillips, E. J. P., și Woodward, J. C. (1996). Substanțe humice ca acceptori de electroni pentru respirația microbiană. Nature 382, 445-448. doi: 10.1038/382445a0
CrossRef Full Text | Google Scholar
Lovley, D. R., Giovannoni, S. J., White, D. C., Champine, J. E., Phillips, E. J., Gorby, Y. A., et al. (1993). Geobacter metallireducens gen. nov. sp. nov., un microorganism capabil să cupleze oxidarea completă a compușilor organici cu reducerea fierului și a altor metale. Arch. Microbiol. 159, 336-344. doi: 10.1007/BF00290916
PubMed Abstract | Text integral | Google Scholar
Lovley, D. R., Holmes, D. E., și Nevin, K. P. (2004). Dissimilatory Fe(III) and Mn(IV) reduction. Adv. Microb. Physiol. 49, 219-286. doi: 10.1111/j.1462-2920.2004.00593.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Marsili, E., Baron, D. B., Shikhare, I. D., Coursolle, D., Gralnick, J. A., Bond, D. R., et al. (2008). Shewanella secretă flavine care mediază transferul extracelular de electroni. Proc. Natl. acad. Sci. U.S.A. 105, 3968-3973. doi: 10.1073/pnas.0710525105
PubMed Abstract | CrossRef Full Text | Google Scholar
McLean, J. S., Majors, P. D., Reardon, C. L., Bilskis, C. L., Reed, S. B., Romine, M. F., et al. (2008). Investigations of structure and metabolism within Shewanella oneidensis MR-1 biofilms. J. Microbiol. Methods 74, 47-56. doi: 10.1016/j.mimet.2008.02.015
PubMed Abstract | CrossRef Full Text | Google Scholar
Meyer, T. E., Tsapin, A. I., Vandenberghe, I., de Smet, L., Frishman, D., Nealson, K. H., et al. (2004). Identificarea a 42 de gene posibile ale citocromului C în genomul Shewanella oneidensis și caracterizarea a șase citocromi solubili. Omics 8, 57-77. doi: 10.1089/153623104773547499
PubMed Abstract | CrossRef Full Text | Google Scholar
Mitchell, A. C., Peterson, L., Reardon, C. L., Reed, S. B., Culley, D. E., Romine, M. R., et al. (2012). Rolul citocromilor de tip c din membrana exterioară MtrC și OmcA în producția, acumularea și detașarea celulelor Shewanella oneidensis MR-1 în timpul respirației pe hematite. Geobiology 10, 355-370. doi: 10.1111/j.1472-4669.2012.00321.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Murray, A. E., Lies, D., Li, G., Nealson, K., Zhou, J., și Tiedje, J. M. (2001). Hibridizarea ADN/ADN la microarrays dezvăluie diferențe specifice genelor între genomurile microbiene strâns înrudite. Proc. Natl. acad. Sci. U.S.A. 98, 98, 9853-9858. doi: 10.1073/pnas.171178898
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., și Myers, J. M. (1997). Clonarea și secvențierea cymA, o genă care codifică un citocrom c tetraheme necesar pentru reducerea fierului(III), fumaratului și nitratului de către Shewanella putrefaciens MR-1. J. Bacteriol. 179, 1143-1152. doi: 10.1128/jb.179.4.1143-1152.1997
PubMed Abstract | CrossRefef Full Text | Google Scholar
Myers, C. R., și Myers, J. M. (2002). MtrB Este necesar pentru încorporarea corectă a citocromilor OmcA și OmcB în membrana externă a Shewanella putrefaciens MR-1. Appl. Environ. Microbiol. 68, 5585-5594. doi: 10.1128/AEM.68.68.11.5585-5594.2002
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., și Nealson, K. H. (1988a). Reducerea manganului bacterian și creșterea cu oxid de mangan ca unic acceptor de electroni. Science 240, 1319-1321. doi: 10.1126/science.240.4857.1319
PubMed Abstract | CrossRef Full Text | Google Scholar
Myers, C. R., and Nealson, K. H. (1988b). Reducerea microbiană a oxizilor de mangan: Interacțiuni cu fierul și sulful. Geochim. Cosmochim. Acta 52, 2727-2732. doi: 10.1016/0016-7037(88)90041-5
CrossRef Full Text | Google Scholar
Myers, J. M., and Myers, C. R. (2001). Rolul pentru citocromii de membrană externă OmcA și OmcB din Shewanella putrefaciens MR-1 în reducerea dioxidului de mangan. Appl. Environ. Microbiol. 67, 260-269. doi: 10.1128/AEM.67.1.260-269.2001
PubMed Abstract | Refef Full Text | Google Scholar
Nealson, K. H., Moser, D. P., și Saffarini, D. A. (1995). Chimiotaxa de acceptor de electroni anaerobi în Shewanella putrefaciens. Appl. Environ. Microbiol. 61, 1551-1554.
PubMed Abstract | Google Scholar
Okamoto, A., Kalathil, S., Deng, X., Hashimoto, K., Nakamura, R., și Nealson, K. H. (2014). Flavinele secretate de celule legate de citocromii membranari dictează reacțiile de transfer de electroni către suprafețe cu sarcină și pH diferite. Sci. Rep. 4:5628. doi: 10.1038/srep05628
PubMed Abstract | Ref. Full Text | Google Scholar
Pirbadian, S., Barchinger, S. E., Leung, K. M., Byun, H. S. S., Jangir, Y., Bouhenni, R. A., et al. (2015). Nanofirele bacteriene din Shewanella oneidensis MR-1 sunt extensii ale membranei externe și periplasmice ale componentelor extracelulare de transport de electroni. Biophys. J. 108:368a. doi: 10.1016/j.bpj.2014.11.2016
CrossRef Full Text | Google Scholar
Reguera, G., McCarthy, K. D., Mehta, T., Nicoll, J. S., Tuominen, M. T., Lovely, D. R., et al. (2005). Transferul extracelular de electroni prin intermediul nanofirelor microbiene. Nature 435, 1098-1101. doi: 10.1038/nature03661
PubMed Abstract | Text integral | Google Scholar
Salas, E. C., Berelson, W. M., Hammond, D. E., Kampf, A. R., și Nealson, K. H. (2010). Impactul tulpinii bacteriene asupra produselor de reducere disimilatorie a fierului. Geochim. Cosmochim. Acta 74, 574-583. doi: 10.1016/j.gca.2009.10.039
PubMed Abstract | CrossRef Full Text | Google Scholar
Saltikov, C. W., Cifuentes, A., Venkateswaran, K., și Newman, D. K. (2003). Sistemul de detoxifiere ars este avantajos, dar nu este necesar pentru respirația As(V) de către tulpina ANA-3 a speciei Shewanella, care poate fi tratată genetic. Appl. Environ. Microbiol. 69, 2800-2809. doi: 10.1128/AEM.69.5.2800-2809.2003
CrossRef Full Text | Google Scholar
Schwalb, C., Chapman, S. K., and Reid, G. A. (2003). Citocromul tetraheme CymA este necesar pentru respirația anaerobă cu dimetil sulfoxid și nitrit în Shewanella oneidensis. Biochemistry 42, 9491-9497. doi: 10.1021/bi034456f
PubMed Abstract | Ref Full Text | Google Scholar
Schwertmann, U., și Cornell, R. M. (2008). Oxizi de fier în laborator. Weinheim: John Wiley & Sons.
Google Scholar
Shi, L., Richardson, D. J., Wang, Z., Kerisit, S. N., Rosso, K. M., Zachara, J. M., et al. (2009). The roles of outer membrane cytochromes of Shewanella and Geobacter in extracellular electron transfer. Environ. Microbiol. Rep. 1, 220-227. doi: 10.1111/j.1758-2229.2009.00035.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Venkateswaran, K., Dollhopf, M. E., Aller, R., Stackebrandt, E., și Nealson, K. H. (1998). Shewanella amazonensis sp. nov., un nou anaerob facultativ reducător de metale din nămolurile de pe platforma amazoniană. Int. J. Syst. Bacteriol. 48, 965-972.
PubMed Abstract | Google Scholar
Venkateswaran, K., Moser, D. P., Dollhopf, M. E., Lies, D. P., Saffarini, D. A., MacGregor, B. J., et al. (1999). Polyphasic taxonomy of the genus Shewanella and description of Shewanella oneidensis sp. nov. Int. J. Syst. Bacteriol. 49, 705-724. doi: 10.1099/00207713-49-2-705
PubMed Abstract | Text integral | Google Scholar
Ziemke, F., Höfle, M. G., Lalucat, J., și Rossellö-Mora, R. (1998). Reclasificarea grupului genomic II al lui Shewanella putrefaciens Owen ca Shewanella baltica sp. nov. Int. J. Syst. Bacteriol. 48, 179-186. doi: 10.1099/00207713-48-1-179
PubMed Abstract | CrossRef Full Text | Google Scholar
.